
古脊椎动物学报 ›› 2023, Vol. 61 ›› Issue (3): 161-181.DOI: 10.19615/j.cnki.2096-9899.230703CSTR: 32090.14.j.cnki.2096-9899.230703
• • 下一篇
冯东昊1,2, 徐光辉1,*( ), 马昕莹3, 任艺1,2
), 马昕莹3, 任艺1,2
收稿日期:2023-03-22
出版日期:2023-07-20
发布日期:2023-07-25
通讯作者:
* xuguanghui@ivpp.ac.cn基金资助:
FENG Dong-Hao1,2, XU Guang-Hui1,*(), MA Xin-Ying3, REN Yi1,2
Received:2023-03-22
Published:2023-07-20
Online:2023-07-25
摘要:
小型近鲱形类(Halecomorphi)贵州中华真颌鱼(Sinoeugnathus kueichowensis)发现于中国云贵地区中三叠世拉丁期海相地层中,曾被归入弓鳍鱼目(Amiiformes)真颌鱼科(Eugnathidae)。基于模式标本和15块新标本的详细观察,对该属种进行了重新描述和分类学修订。对这些标本的研究揭示了该属种在早前研究中一些未被描述或识别错误的形态特征,包括斧状的眶前骨,两个宽大的次眶骨,上颌骨具有感觉管和三对额外肩胛骨。首次将中华真颌鱼纳入近鲱形类分支系统学研究中;结果显示,中华真颌鱼与云南罗平安尼期的复兴鱼(Subortichthys)构成姐妹群,两者与欧洲圣乔治山地区拉丁期的Allolepidotus和始真颌鱼(Eoeugnathus)一起组成预言鱼目(Ionoscopiformes)底部的一个单系群(命名为复兴鱼科)。中华真颌鱼的分类学修订为了解预言鱼目的系统发育关系和古地理演化提供了新的信息。
中图分类号:
冯东昊, 徐光辉, 马昕莹, 任艺. 云贵地区中三叠世近鲱形类贵州中华真颌鱼(Sinoeugnathus kueichowensis)的分类学修订. 古脊椎动物学报, 2023, 61(3): 161-181.
FENG Dong-Hao, XU Guang-Hui, MA Xin-Ying, REN Yi. Taxonomic revision of Sinoeugnathus kueichowensis (Halecomorphi, Holostei) from the Middle Triassic of Guizhou and Yunnan, China. Vertebrata Palasiatica, 2023, 61(3): 161-181.

Fig. 1 Holotype, two new specimens and reconstruction of Sinoeugnathus kueichowensis A. IVPP V2433 (holotype); B. V24315; C. V30784; D. reconstruction
| Specimen | SL | HL | BD | TL | PVL | PDL | PAL |
|---|---|---|---|---|---|---|---|
| V2433 | 53.00 | 19.35 | 21.51 | 66.41 | 29.79 | - | 40.40 |
| V19007 | 66.20 | 20.88 | 23.16 | 83.28 | - | 40.16 | 52.24 |
| V24315 | 64.22 | 20.94 | 26.92 | 82.56 | 35.18 | 41.70 | 49.66 |
| V24316 | 52.20 | 18.98 | 22.48 | 63.68 | 29.00 | 29.28 | 40.32 |
| V24317 | 58.66 | 20.40 | 22.24 | 73.06 | 33.92 | 37.90 | 45.10 |
| V24320 | 65.40 | 22.74 | 25.00 | 83.06 | - | 40.14 | 49.24 |
| V30784 | 69.58 | 22.12 | 23.42 | 87.54 | 37.44 | 39.52 | 52.24 |
| V30786 | 59.68 | 19.06 | 22.86 | 70.78 | 34.16 | 37.46 | 46.38 |
| V30787 | 61.32 | 21.18 | 22.36 | 69.12 | 31.52 | 35.98 | 44.14 |
| V30789 | 60.50 | 22.04 | 21.80 | 75.66 | 31.32 | - | 44.46 |
| V30790 | 56.56 | 21.44 | 21.72 | 74.88 | 30.50 | 37.52 | 42.92 |
Table 1 Measurement data of 11 specimens of Sinoeugnathus kueichowensis (mm)
| Specimen | SL | HL | BD | TL | PVL | PDL | PAL |
|---|---|---|---|---|---|---|---|
| V2433 | 53.00 | 19.35 | 21.51 | 66.41 | 29.79 | - | 40.40 |
| V19007 | 66.20 | 20.88 | 23.16 | 83.28 | - | 40.16 | 52.24 |
| V24315 | 64.22 | 20.94 | 26.92 | 82.56 | 35.18 | 41.70 | 49.66 |
| V24316 | 52.20 | 18.98 | 22.48 | 63.68 | 29.00 | 29.28 | 40.32 |
| V24317 | 58.66 | 20.40 | 22.24 | 73.06 | 33.92 | 37.90 | 45.10 |
| V24320 | 65.40 | 22.74 | 25.00 | 83.06 | - | 40.14 | 49.24 |
| V30784 | 69.58 | 22.12 | 23.42 | 87.54 | 37.44 | 39.52 | 52.24 |
| V30786 | 59.68 | 19.06 | 22.86 | 70.78 | 34.16 | 37.46 | 46.38 |
| V30787 | 61.32 | 21.18 | 22.36 | 69.12 | 31.52 | 35.98 | 44.14 |
| V30789 | 60.50 | 22.04 | 21.80 | 75.66 | 31.32 | - | 44.46 |
| V30790 | 56.56 | 21.44 | 21.72 | 74.88 | 30.50 | 37.52 | 42.92 |
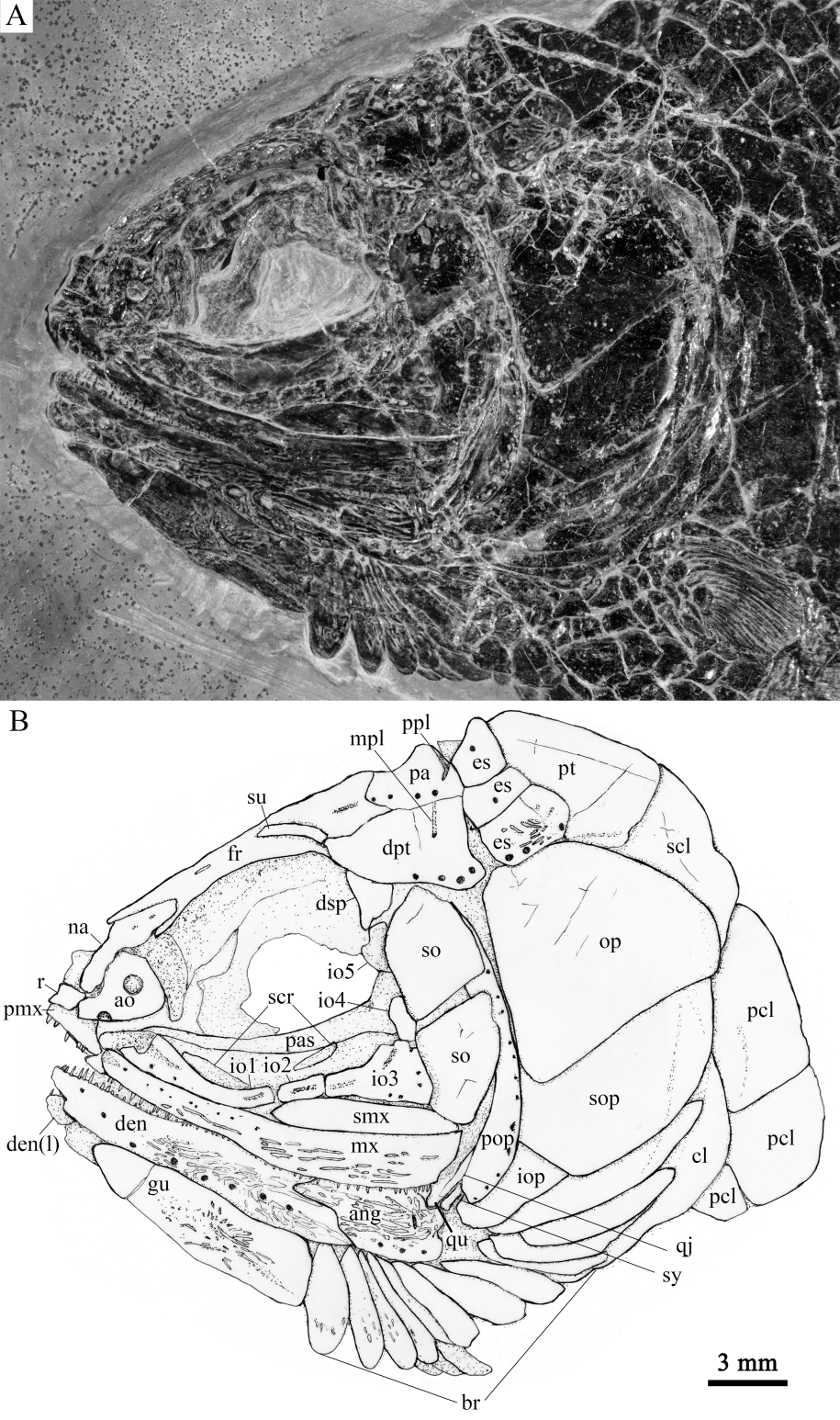
Fig. 2 Photograph (A) and line-drawing (B) of skull and pectoral girdle of Sinoeugnathus kueichowensis, IVPP V24315
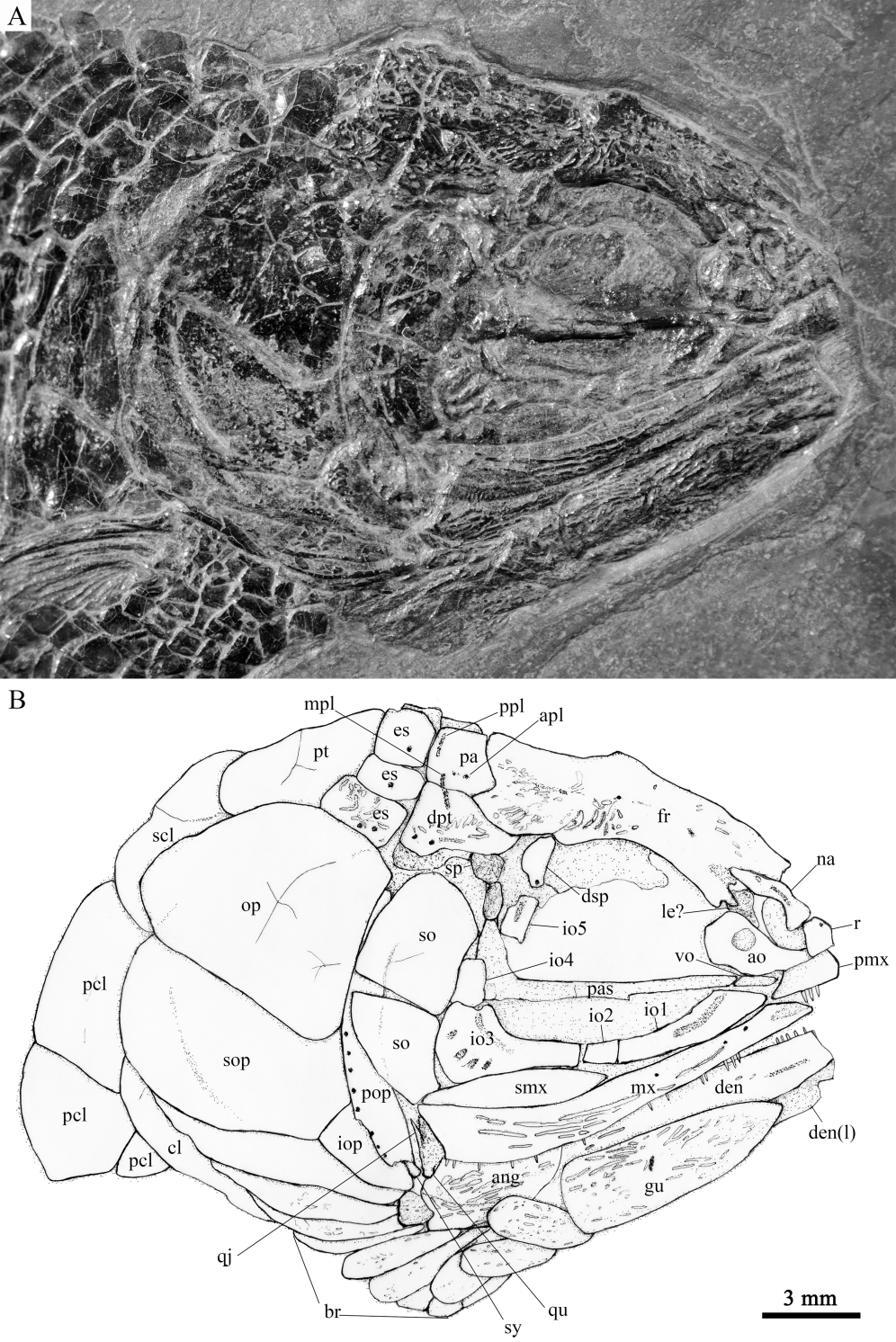
Fig. 3 Photograph (A) and line-drawing (B) of skull and pectoral girdle of Sinoeugnathus kueichowensis, IVPP V30784
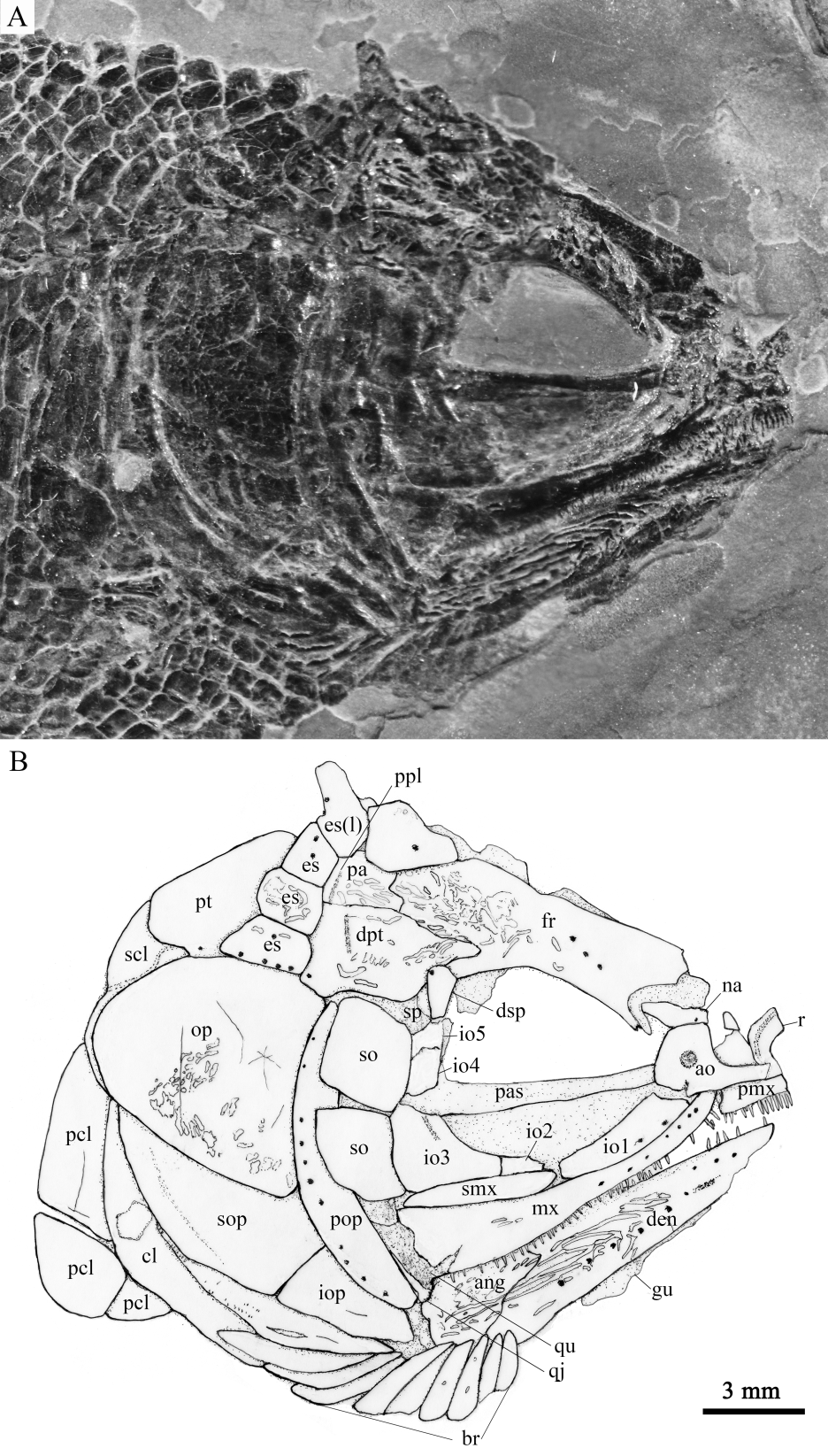
Fig. 4 Photograph (A) and line-drawing (B) of skull and pectoral girdle of Sinoeugnathus kueichowensis, IVPP V30785
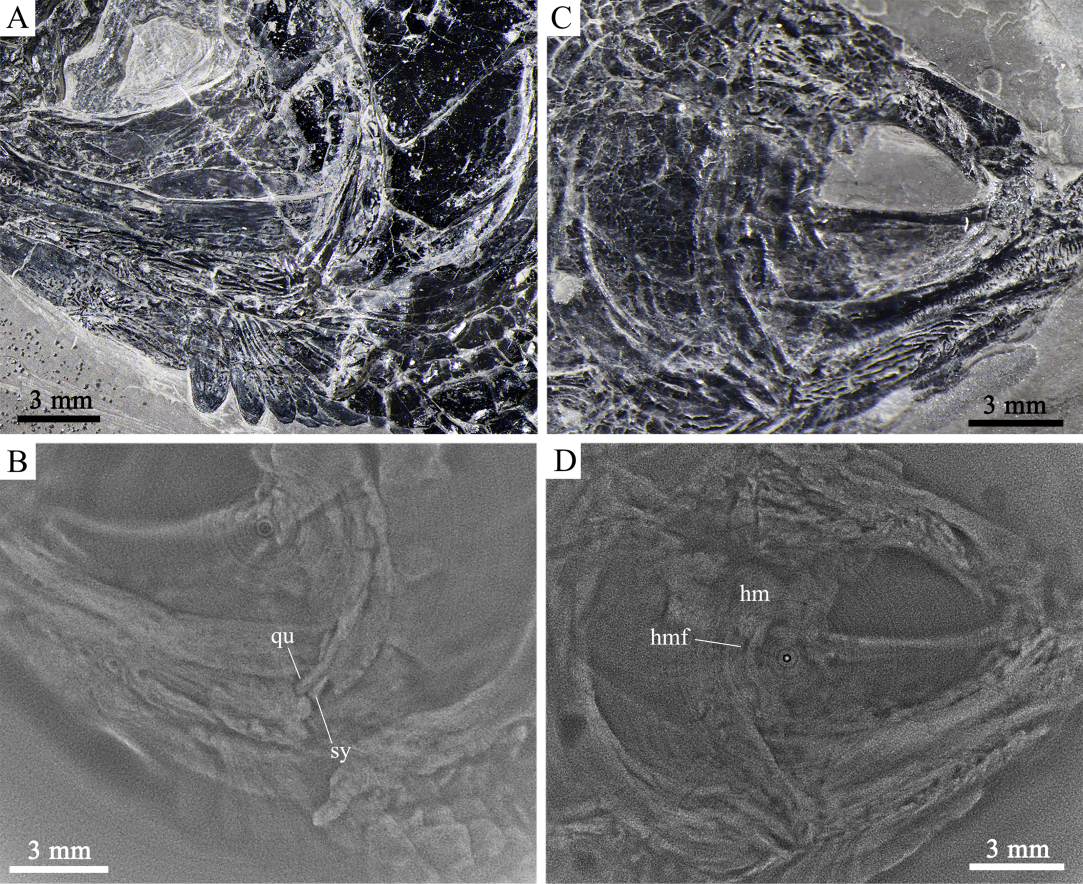
Fig. 5 Cranial bones of Sinoeugnathus kueichowensis A. IVPP V24315; B. X-ray computed tomography of A, showing both the quadrate symplectic condyles for the jaw hinge; C. V30785; D. X-ray computed tomography of C, showing the hyomandibula and the foramen for the hyomandibular trunk of facial nerve (VII)
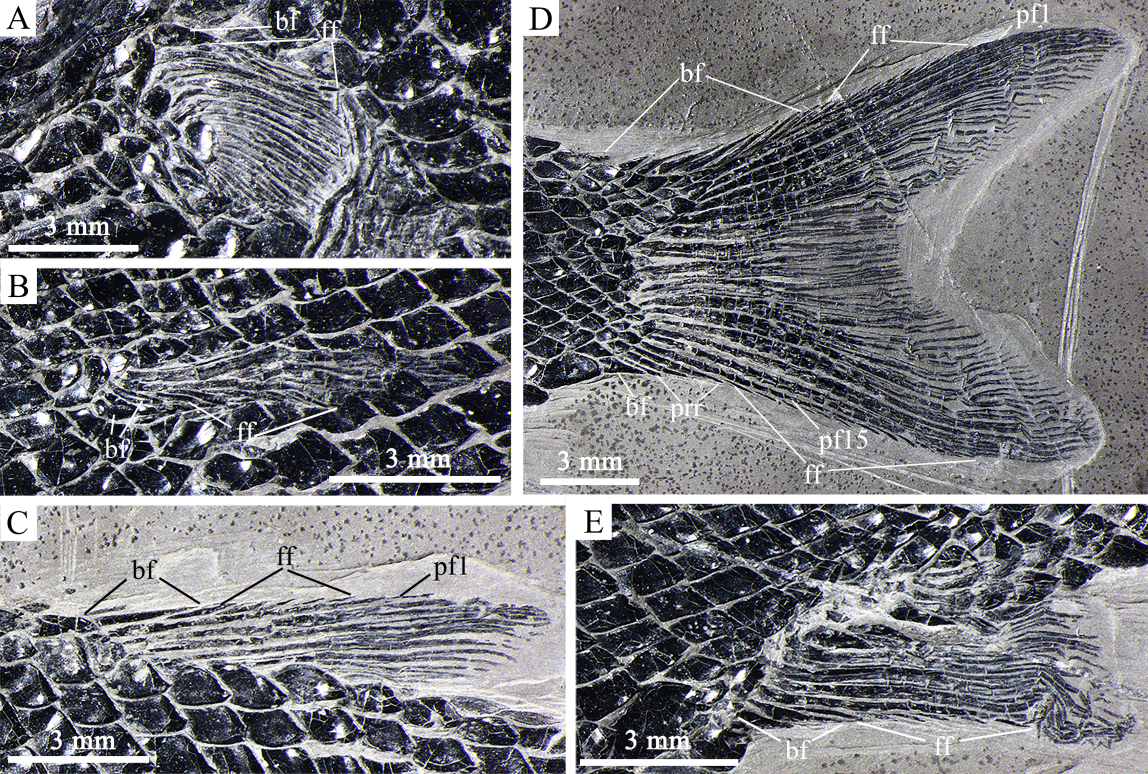
Fig. 6 Fins of Sinoeugnathus kueichowensis, IVPP V24315 A. left pectoral fin; B. left pelvic fin; C. dorsal fin; D. caudal fin; E. anal fin

Fig. 7 Complete specimen and lateral line scales of Sinoeugnathus kueichowensis, IVPP V30785 A. complete specimen; B. scales in predorsal region with arrow showing pore of additional lateral line; C. main lateral line scales in posterior flank region with arrows showing pores of lateral line
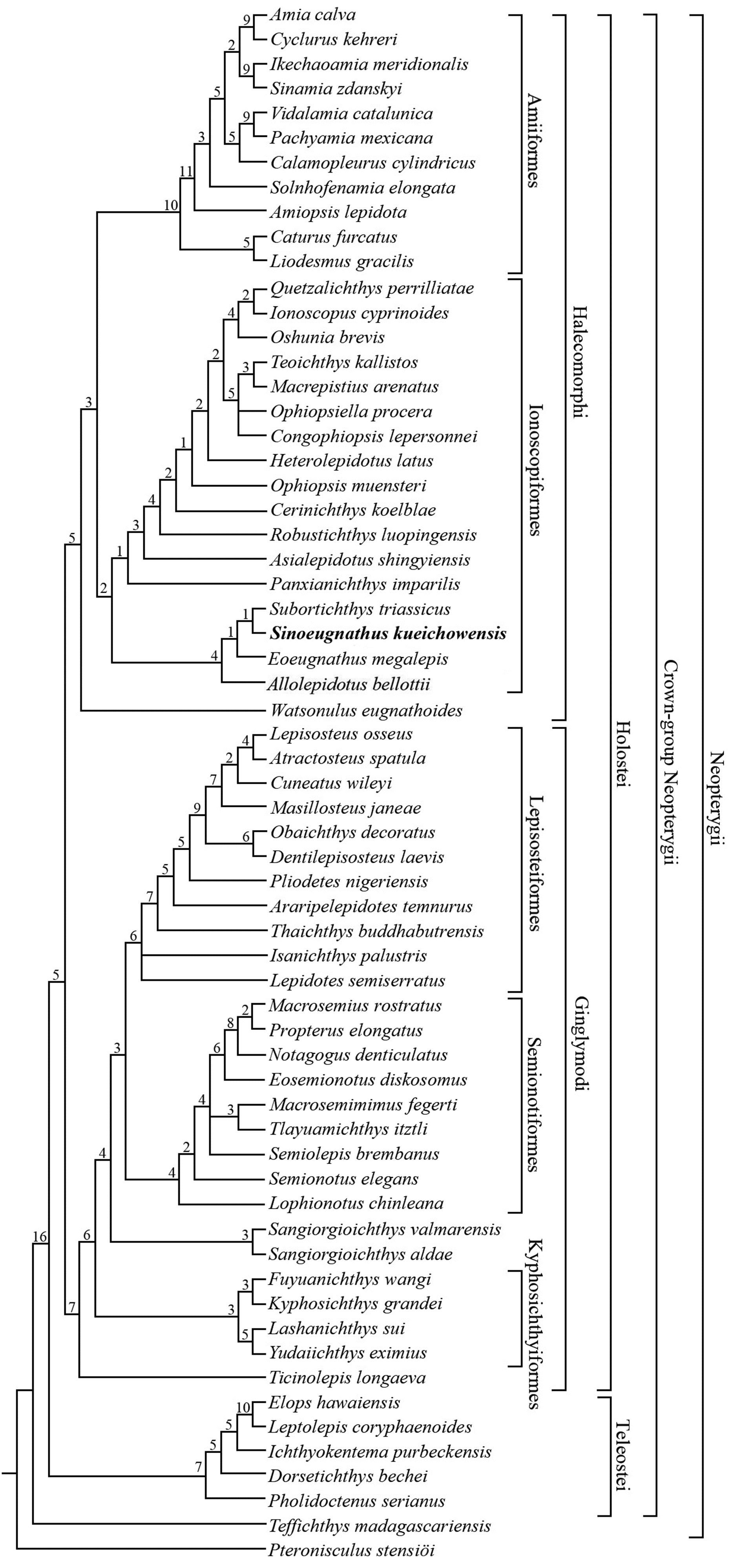
Fig. 8 Strict consensus of 48 most parsimonious trees illustrating the phylogenetic relationships of Sinoeugnathus within the Neopterygii Tree length = 661 steps; consistency index (CI) = 0.4115; retention index (RI) = 0.7665 Digits above nodes indicate Bremer decay indices. For character descriptions and codings for the sampled taxa, see the online supplementary material
| Species | MXL | MDL | HL | MXL/MDL | MDL/HL | SL | Specimen |
|---|---|---|---|---|---|---|---|
| Sinoeugnathus kueichowensis | 13 | 15 | 22 | 0.87 | 0.68 | 69.6 | IVPP V30784 |
| Subortichthys triassicus | 17 | 19 | 28 | 0.89 | 0.68 | 72 | IVPP V20680 |
| Allolepidotus bellotti | 15.5 | 16.5 | 26 | 0.94 | 0.63 | 85 | MCSNIO P647 |
| Eoeugnathus megalepis | 21 | 25 | 37 | 0.84 | 0.68 | 98 | PIMUZ A/I 1964 |
| Panxianichthys imparilis | 20 | 27 | 50 | 0.74 | 0.54 | 159 | IVPP V19971 |
| Aisalepidotus shingyiensis | 36 | 48 | 91 | 0.75 | 0.53 | 273 | IVPP V23010 |
| Robustichthys luopingensis | 14 | 24 | 45 | 0.62 | 0.54 | 140 | IVPP V18568 |
| Robustichthys luopingensis | - | 53 | 100 | - | 0.53 | 360 | IVPP V20414 |
Table 2 Measurements for eight specimens of Triassic ionoscopiforms (mm)
| Species | MXL | MDL | HL | MXL/MDL | MDL/HL | SL | Specimen |
|---|---|---|---|---|---|---|---|
| Sinoeugnathus kueichowensis | 13 | 15 | 22 | 0.87 | 0.68 | 69.6 | IVPP V30784 |
| Subortichthys triassicus | 17 | 19 | 28 | 0.89 | 0.68 | 72 | IVPP V20680 |
| Allolepidotus bellotti | 15.5 | 16.5 | 26 | 0.94 | 0.63 | 85 | MCSNIO P647 |
| Eoeugnathus megalepis | 21 | 25 | 37 | 0.84 | 0.68 | 98 | PIMUZ A/I 1964 |
| Panxianichthys imparilis | 20 | 27 | 50 | 0.74 | 0.54 | 159 | IVPP V19971 |
| Aisalepidotus shingyiensis | 36 | 48 | 91 | 0.75 | 0.53 | 273 | IVPP V23010 |
| Robustichthys luopingensis | 14 | 24 | 45 | 0.62 | 0.54 | 140 | IVPP V18568 |
| Robustichthys luopingensis | - | 53 | 100 | - | 0.53 | 360 | IVPP V20414 |
| [1] |
Benton M J, Zhang Q Y, Hu S X et al., 2013. Exceptional vertebrate biotas from the Triassic of China, and the expansion of marine ecosystems after the Permo-Triassic mass extinction. Earth Sci Rev, 125: 199-243
DOI URL |
| [2] | Brough J, 1939. The Triassic fishes of Besano, Lombardy. London: British Museum (Natural History). 1-117 |
| [3] | Bürgin T, 2004. Eosemionotus ceresiensis sp. nov., a new semionotiform fish (Actinopterygii, Halecostomi) from the Middle Triassic of Monte San Giorgio (Southern Switzerland). In: Arratia G, Tintori A eds. Mesozoic Fishes 3-Systematics, Paleoenvironments and Biodiversity 3. 239-251 |
| [4] |
Cavin L, 2010. Diversity of Mesozoic semionotiform fishes and the origin of gars (Lepisosteidae). Naturwissenschaften, 97: 1035-1040
DOI PMID |
| [5] |
Ebert M, 2018. Cerinichthys koelblae, gen. et sp. nov., from the Upper Jurassic of Cerin, France, and its phylogenetic setting, leading to a reassessment of the phylogenetic relationships of Halecomorphi (Actinopterygii). J Vert Paleont, 38: e1420071
DOI URL |
| [6] | Folkvord A, Hunter J R, 1986. Size-specific vulnerability of northern anchovy, Engraulis mordax, larvae to predation by fishes. Fish Bull, 84: 859-869 |
| [7] |
Friedman M, 2015. The early evolution of ray-finned fishes. Paleontology, 58: 213-228
DOI URL |
| [8] | Gardiner B G, 1960. A revision of certain actinopterygian and coelacanth fishes, chiefly from the Lower Lias. Bull Br Mus Nat Hist Geol, 4(7): 241-384 |
| [9] | Geng B H, Zhu M, Jin F, 2009. A revision and phylogenetic analysis of Guizhoucoelacanthus (Sarcopterygii, Actinistia) from the Triassic of China. Vert PalAsiat, 47: 165-177 |
| [10] |
Gill E L, 1923. The Permian fishes of the genus Acentrophorus. Proc Zool Soc Lond, 93: 19-40
DOI URL |
| [11] | Grande L, 2010. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. The resurrection of holostei. Copeia, 10(Suppl): 1-871 |
| [12] | Grande L, Bemis W E, 1998. A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy: an empirical search for interconnected patterns of natural history. Soc Vert Paleont Mem (Suppl J Vert Paleont), 4: 1-690 |
| [13] | Herzog A, 2003. Eine Neubeschreibung der Gattung Eoeugnathus, Brough, 1939 (Actinopterygii; Halecomorphi) aus der alpinen Mitteltrias Graubündens. Palaönt Zeit, 77(1): 223-240 |
| [14] |
Hurley I A, Mueller R L, Dunn K A et al., 2007. A new time-scale for ray-finned fish evolution. Proc R Soc B, 274: 489-498
DOI URL |
| [15] |
Jiang D Y, Motani R, Tintori A et al., 2020. Evidence supporting predation of 4-m marine reptile by Triassic megapredator. iScience, 23: 101347
DOI URL |
| [16] | Jin F, 2001. Notes on the discovery of Birgeria in China. Vert PalAsiat, 39: 168-176 |
| [17] |
Karpouzi V S, Stergiou K I, 2003. The relationships between mouth size and shape and body length for 18 species of marine fishes and their trophic implications. J Fish Biol, 62: 1353-1365
DOI URL |
| [18] |
Li C, Wu X C, Zhao L J et al., 2016. A new armored archosauriform (Diapsida: Archosauromorpha) from the marine Middle Triassic of China, with implications for the diverse life styles of archosauriforms prior to the diversification of Archosauria. Sci Nat, 103: 95
DOI URL |
| [19] | Li J L, 2006. A brief summary of the Triassic marine reptiles of China. Vert PalAsiat, 44: 99-108 |
| [20] | Li Z G, Sun Z Y, Jiang D Y et al., 2016. LA-ICP-MS Zircon U-Pb age of the fossil layer of Triassic Xingyi Fauna from Xingyi, Guizhou, and its significance. Geol Rev, 62: 779-790 |
| [21] | Liu G B, Yin G Z, Wang X H, 2002. On the most primitive amiid fish from Upper Triassic of Xingyi, Guizhou. Acta Palaeont Sin, 41: 461-463 |
| [22] | Liu G B, Yin G Z, Wang X H et al., 2003. New discovered fishes from Keichousaurus bearing horizon of Late Triassic in Xingyi of Guizhou. Acta Palaeont Sin, 42: 346-366 |
| [23] | Lombardo C, 2001. Actinopterygians from the Middle Triassic of northern Italy and Canton Ticino (Switzerland): anatomical descriptions and nomenclatural problems. Riv Ital Paleont Stratigr, 107: 345-369 |
| [24] |
López-Arbarello A, Ebert M, 2023. Taxonomic status of the caturid genera (Halecomorphi, Caturidae) and their Late Jurassic species. R Soc Open Sci, 10: 221318
DOI URL |
| [25] |
López-Arbarello A, Sferco E, 2018. Neopterygian phylogeny: the mergerassay. R Soc Open Sci, 5: 172337
DOI URL |
| [26] |
López-Arbarello A, Sun Z Y, Sferco E et al., 2011. New species of Sangiorgioichthys Tintori and Lombardo, 2007 (Neopterygii, Semionotiformes) from the Anisian of Luoping (Yunnan Province, South China). Zootaxa, 2749: 25-39
DOI URL |
| [27] | Ma X Y, Xu G H, 2017. A new ionoscopiform fish (Holostei: Halecomorphi, Ionoscopiformes) from the Middle Triassic (Anisian) of Yunnan, China. Vert PalAsiat, 55: 92-106 |
| [28] |
Magnhagen C, Heibo E, 2001. Gape size allometry in pike reflects variation between lakes in prey availability and relative body depth. Funct Ecol, 15: 754-762
DOI URL |
| [29] | Metcalfe I, 2011. Palaeozoic-Mesozoic history of SE Asia. In: Hall R, Cottam M A, Wilson M E J eds. The SE Asian Gateway: History and Tectonics of the Australia-Asia Collision. Geol Soc, Spec Publ, London, 355: 7-35 |
| [30] |
Near T J, Eytan R I, Dornburg A et al., 2012. Resolution of ray-finned fish phylogeny and timing of diversification. Proc Nat Acad Sci, 109: 13698-13703
DOI URL |
| [31] |
Olsen P E, 1984. The skull and pectoral girdle of the parasemionotid fish Watsonulus eugnathoides from the Early Triassic Sakamena group of Madagascar, with comments on the relationships of the holostean fishes. J Vert Paleont, 4: 481-499
DOI URL |
| [32] | Patterson C, 1973. Interrelationships of holosteans. In: Greenwood P H, Miles R S, Patterson C eds. Interrelationships of Fishes. Zool J Linn Soc, 53(Suppl): 233-305 |
| [33] |
Scharf S F, Juanes F, Rountree R A, 2000. Predator size-prey size relationships of marine fish predators: interspecific variation and effects of ontogeny and body size on trophic-niche breadth. Mar Ecol Prog Ser, 208: 229-248
DOI URL |
| [34] |
Shang Q H, Wu X C, Li C, 2020. A new Ladinian nothosauroid (Sauropterygia) from Fuyuan, Yunnan Province, China. J Vert Paleont, 40: e1789651
DOI URL |
| [35] | Su D Z, 1959. Triassic fishes from Kueichow, Southwest China. Vert PalAsiat, 3: 205-210 |
| [36] |
Sun Z Y, Tintori A, Xu Y Z et al., 2016. A new non-parasemionotiform order of the Halecomorphi (Neopterygii, Actinopterygii) from the Middle Triassic of Tethys. J Syst Palaeont, 15: 223-240
DOI URL |
| [37] | Swofford D L, 2003. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0b10. Sunderland, Massachusetts: Sinauer Associates |
| [38] | Tintori A, Lombardo C, 2007. A new early Semionotidae (Semionotiformes, Actinopterygii) from the Upper Ladinian of the Monte San Giorgio area (southern Switzerland and northern Italy). Riv Ital Paleont Stratigr, 113: 369-381 |
| [39] |
Tintori A, Hitij T, Jiang D Y et al., 2014. Triassic actinopterygian fishes: the recovery after the end-Permian crisis. Integr Zool, 9: 349-411
DOI URL |
| [40] | Tintori A, Sun Z Y, Lombardo C et al., 2015. Oldest stem Teleostei from the late Ladinian (Middle Triassic) of southern China. Riv Ital Paleont Stratigr, 121: 285-296 |
| [41] | Westoll T S, 1944. The Haplolepidae, a new family of Late Carboniferous bony fishes - a study in taxonomy and evolution. Bull Am Mus Nat Hist, 83: 1-121 |
| [42] |
Xu G H, 2019. Osteology and phylogeny of Robustichthys luopingensis, the largest holostean fish in the Middle Triassic. PeerJ, 7: e7184
DOI URL |
| [43] | Xu G H, 2020. A new species of Luganoia (Luganoiidae, Neopterygii) from the Middle Triassic Xingyi Biota, Guizhou, China. Vert PalAsiat, 58: 267-282 |
| [44] |
Xu G H, Ma X Y, 2018. Redescription and phylogenetic reassessment of Asialepidotus shinyiensis (Holostei: Halecmorphi) from the Middle Triassic (Ladinian) of China. Zool J Linn Soc, 184(1): 95-114
DOI URL |
| [45] | Xu G H, Shen C C, 2015. Panxianichthys imparilis gen. et sp. nov., a new ionoscopiform (Halecomorphi) from the Middle Triassic of Guizhou, China. Vert PalAsiat, 53: 1-15 |
| [46] |
Xu G H, Wu F X, 2012. A deep-bodied ginglymodian fish from the Middle Triassic of eastern Yunnan Province, China, and the phylogeny of lower neopterygians. Chinese Sci Bull, 57: 111-118
DOI URL |
| [47] |
Xu G H, Zhao L J, Gao K Q et al., 2012. A new stem-neopterygian fish from the Middle Triassic of China shows the earliest over-water gliding strategy of the vertebrates. Proc R Soc B, 280: 20122261
DOI URL |
| [48] |
Xu G H, Zhao L J, Coates M I, 2014. The oldest ionoscopiform from China sheds new light on the early evolution of halecomorph fishes. Biol Lett, 10: 20140204
DOI URL |
| [49] |
Xu G H, Gao K Q, Coates M I, 2015. Taxonomic revision of Plesiofuro mingshuica from the Lower Triassic of northern Gansu, China, and the relationships of early neopterygian clades. J Vert Paleont, 35(6): e1001515
DOI URL |
| [50] |
Xu G H, Ma X Y, Ren Y, 2018. Fuyuanichthys wangi gen. et sp. nov. from the Middle Triassic (Ladinian) of China highlights the early diversification of ginglymodian fishes. PeerJ, 6: e6054
DOI URL |
| [51] | Young C C, 1958. On the new pachypleurosauroidea from Keichow, South-West China. Vert PalAsiat, 2: 69-81 |
| [52] | Zou X D, Balini M, Jiang D Y et al., 2015. Ammonoids from the Zhuganpo Member of the Falang Formation at Nimaigu and their relevance for dating the Xingyi Fossil-Lagerstaette (late Ladinian, Guizhou, China). Riv Ital Paleont Stratigr, 121: 135-161 |
| [1] | 徐光辉, 袁志伟, 任艺, 廖浚伶, 赵丽君, 宋海军. 江苏安徽早三叠世裂齿鱼科一新种:吴氏三叠鱼. 古脊椎动物学报, 2024, 62(3): 165-185. |
| [2] | 石雨泰, 刘俊. 博格达吐鲁番兽(二齿兽类)的骨骼学. 古脊椎动物学报, 2024, 62(3): 186-200. |
| [3] | 王维, 雷洪, 李淳. 云南中三叠世一恐头龙科小型主龙型类. 古脊椎动物学报, 2024, 62(1): 13-32. |
| [4] | 薛钦元, 余逸伦, 潘照晖, 朱幼安, 朱敏. 节甲鱼类(有颌脊椎动物干群)系统发育多样性的衰落与泥盆纪重大环境-生物事件耦合. 古脊椎动物学报, 2024, 62(1): 1-12. |
| [5] | 罗彦超, 朱敏, 卢立伍, 潘照晖. 湖南中泥盆统跳马涧组中华沟鳞鱼再研究. 古脊椎动物学报, 2023, 61(4): 261-276. |
| [6] | 尚庆华, 李淳, 王维. 云南罗平中三叠世安尼期鳍龙类Nothosaurus一新种. 古脊椎动物学报, 2022, 60(4): 249-270. |
| [7] | 刘俊. 鄂尔多斯盆地陕北肯氏兽组合带的肯氏兽型二齿兽类. 古脊椎动物学报, 2022, 60(3): 212-248. |
| [8] | 武瑞, 涂黎, 韩凤禄. 陕西鄂尔多斯盆地中三叠统一离片椎类牙齿化石. 古脊椎动物学报, 2022, 60(1): 54-58. |
| [9] | 任艺, 徐光辉. 云南罗平中三叠世翼鳕属一新种及早期辐鳍鱼类系统发育关系. 古脊椎动物学报, 2021, 59(3): 169-199. |
| [10] | 肖博, 盛桂莲, 袁俊霞, 王斯人, 胡家铭, 陈顺港, 姬海龙, 侯新东, 赖旭龙. 中国东北鹿亚科动物亚化石的古DNA分子鉴定及系统发育分析. 古脊椎动物学报, 2020, 58(4): 328-337. |
| [11] | 徐光辉. 贵州中三叠世兴义生物群卢加诺鱼属(卢加诺鱼科,新鳍鱼类)一新种. 古脊椎动物学报, 2020, 58(4): 267-282. |
| [12] | 张驰. MrBayes分子钟定年之程序. 古脊椎动物学报, 2019, 57(3): 241-252. |
| [13] | 徐光辉, 马昕莹, 吴飞翔, 任艺. 云南中三叠世拱鱼目一新属种及早期铰齿鱼类系统发育关系再评估. 古脊椎动物学报, 2019, 57(3): 181-204. |
| [14] | 廖俊棋, 徐星. 意外北票龙(兽脚类:镰刀龙类)头部骨骼学研究. 古脊椎动物学报, 2019, 57(2): 117-132. |
| [15] | 王敏, 邹晶梅, 周忠和. 孔子鸟目(鸟类:尾综骨类)的分类厘定. 古脊椎动物学报, 2019, 57(1): 1-37. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||