
古脊椎动物学报 ›› 2021, Vol. 59 ›› Issue (2): 138-168.DOI: 10.19615/j.cnki.1000-3118.210325
• • 上一篇
安晓青1,2,3( ), 张兆群1,4,5
), 张兆群1,4,5
收稿日期:2020-10-03
出版日期:2021-05-20
发布日期:2021-04-20
通讯作者:
angelone@uniroma3.it基金资助:
Chiara ANGELONE1,2,3(), ZHANG Zhao-Qun1,4,5
Received:2020-10-03
Published:2021-05-20
Online:2021-04-20
摘要:
早渐新世是新生代全球气候变化的关键时期。这一时期发生了重大的哺乳动物群转换事件,在欧洲称之为 “大间断”, 在亚洲则为 “蒙古重建”。然而,关于渐新世哺乳动物各谱系的形态变化尚缺少详细探讨。兔形目是现代哺乳动物中多样性最低的类群之一,在全球的化石记录中很常见,尤其是在亚洲。渐新世时,许多古老的始新世兔形类灭绝并被更进步的类群所取代。内蒙古的新发现及对老标本的再研究使我们得以修订鄂尔多斯兔(Ordolagus) (亚洲常见的兔形类,可能源自中-晚始新世的戈壁兔Gobiolagus)。在内蒙古,我们识别出Ordolagus在早渐新世初期就已存在。与亚洲和北美的同期或稍早的兔形类的对比表明,Ordolagus具有一些显著的牙齿形态特征(p3下前边尖发达,颊齿完全高冠,p4-m2下三角座和下跟座舌侧相连等), 这些也是现生兔形类的主要特征。Ordolagus中这些特征的出现与重大全球或区域气候变化同步。对亚洲早期兔形类的进一步研究,并参考其他小哺乳动物与区域气候因素的研究,将有助于完善兔形类作为古气候代用指标的作用。
中图分类号:
安晓青, 张兆群. 气候变化与早期兔形类的演化:基于内蒙古Ordolagus新材料的研究. 古脊椎动物学报, 2021, 59(2): 138-168.
Chiara ANGELONE, ZHANG Zhao-Qun. Climate change and evolution of early lagomorphs (Mammalia): a study perspective based on new materials of Ordolagus from Nei Mongol (northern China). Vertebrata Palasiatica, 2021, 59(2): 138-168.
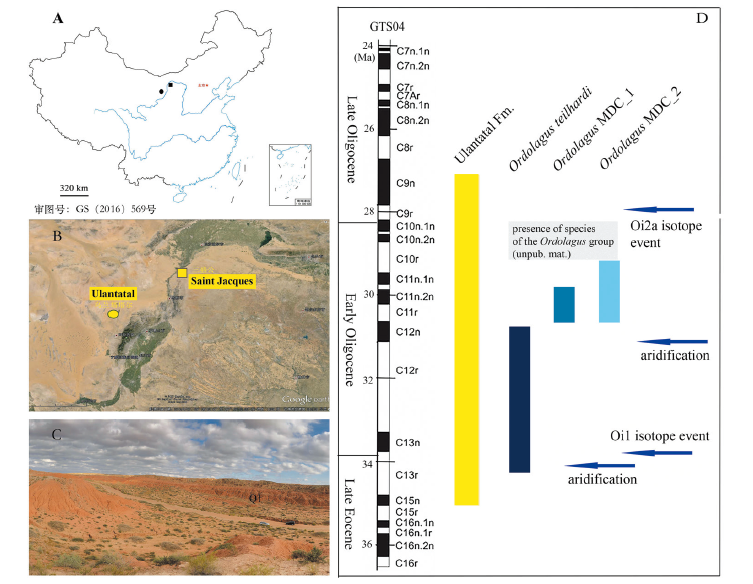
Fig. 1 Geographical location of the fossil sites and temporal distribution of the taxa addressed in this paper relative to the Ulantatal Fm (Nei Mongol, northern China) A. location of Ulantatal and Saint Jacques; B. detail of location of Ulantatal and Saint Jacques (Google maps); C. landscape with fossil horizon Q1 in Saint Jacques; D. estimated chronological distribution of Ordolagus from Ulantatal and Saint Jacques (see Appendix I for details about the horizons and sections and their age estimation); the shaded area represents the chronological distribution of the genus Ordolagus basing on materials not yet studied in detail (ZZQ, IVPP); palaeomagnetic data from Wasiljeff et al. (2020); climatic data (blue arrows) from Sun and Windley (2015), Hauptvogel et al. (2017), Richoz et al. (2017), Ao et al. (2020) with references
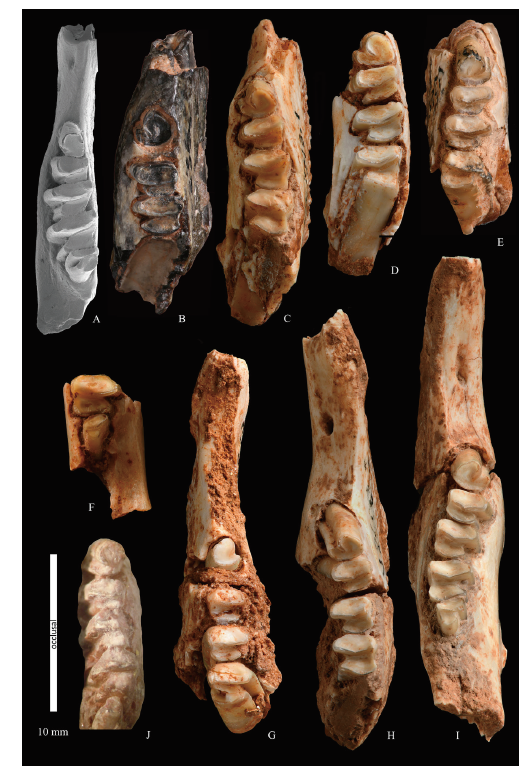
Fig. 2 Gobiolagus tolmachovi and Ordolagus teilhardi from Nei Mongol, China A. Gobiolagus tolmachovi: left lower jaw, AMNH 26142, Nei Mongol, late Middle Eocene (Burke, 1941; Meng et al., 2005); B-J. Ordolagus teilhardi: B. left lower jaw, holotype, AMNH 20236;C. left lower jaw, IVPP V 26013.1; D. left lower jaw, V 26075.1; E. right lower jaw, V 26014; F. part of right lower jaw, V 26013.2; G. right lower jaw (juvenile), V 26017; H. left lower jaw, V 26012.2; I. right lower jaw, V 26012.1; J. left lower jaw, former paratype of O. teilhardi (not included in this review, MNHN, without formal inventory number; see Teilhard de Chardin, 1926:26, fig. 14C)
| p3 | p4 | m1 | m2 | m3 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | W | L/W | L | W | Wtal | L | W | Wtal | L | W | Wtal | L | W | Wtal | ||||||
| Ordolagus teilhardi | AMNH 20236 | 2.05 | 2.27 | 0.90 | 2.08 | 2.77 | 2.02 | 2.37 | 2.91 | 2.27 | ||||||||||
| IVPP V 26012.1 | 1.93 | 2.35 | 0.82 | 1.93 | 2.60 | 2.05 | 2.25 | 2.80 | 2.00 | 2.50 | 2.88 | 2.33 | 1.58 | 1.90 | 1.05 | |||||
| V 26012.2 | 1.87 | 2.36 | 0.79 | 2.05 | 2.72 | 2.00 | ||||||||||||||
| V 26013.1 | 1.90 | 2.33 | 0.82 | 2.03 | 2.68 | 1.93 | 2.55 | 3.18 | 2.15 | 2.75 | 3.10 | 2.50 | ||||||||
| V 26013.2 | 2.05 | 2.29 | 0.90 | 2.05 | 2.43 | 1.76 | 2.55 | 2.91 | 2.12 | 1.10 | 1.62 | 0.83 | ||||||||
| V 26014 | 2.16 | 2.40 | 0.90 | 1.85 | 2.70 | 1.90 | 2.23 | 3.06 | 1.91 | 2.20 | 3.12 | |||||||||
| V 26015.1 | 1.88 | 2.20 | 0.85 | |||||||||||||||||
| V 26015.2 | 1.80 | 2.75 | 2.05 | |||||||||||||||||
| V 26015.3 | 2.20 | 2.85 | 2.13 | |||||||||||||||||
| V 26017 | 1.57 | 1.71 | 0.92 | |||||||||||||||||
| V 26017* | 2.02* | 2.31* | 0.88 | |||||||||||||||||
| V 26075.1 | 1.93 | 2.46 | 0.78 | 1.98 | 2.73 | 2.22 | ||||||||||||||
| V 26075.3 | 1.88* | |||||||||||||||||||
| V 26075.4 | 2.10* | |||||||||||||||||||
| V 26075.5 | 1.76* | |||||||||||||||||||
| N | 13 | 10 | 10 | 8 | 8 | 8 | 5 | 5 | 5 | 4 | 4 | 3 | 2 | 2 | 2 | |||||
| Max | 2.16 | 2.46 | 0.92 | 2.08 | 2.77 | 2.22 | 2.55 | 3.18 | 2.27 | 2.75 | 3.12 | 2.50 | 1.58 | 1.90 | 1.05 | |||||
| Min | 1.57 | 1.71 | 0.78 | 1.80 | 2.43 | 1.76 | 2.20 | 2.80 | 1.91 | 2.20 | 2.88 | 2.12 | 1.10 | 1.62 | 0.83 | |||||
| Mean | 1.93 | 2.27 | 0.86 | 1.97 | 2.67 | 1.99 | 2.32 | 2.96 | 2.09 | 2.50 | 3.00 | 2.31 | 1.34 | 1.76 | 0.94 | |||||
| OrdolagusMCD_1 | V 6268.13 | 2.25 | 2.13 | 1.06 | 1.93 | 2.87 | 2.05 | 2.23 | 3.17 | 1.98 | 2.52 | 3.22 | 2.30 | |||||||
| V 26018.1 | 1.90 | 1.86 | 1.03 | 1.62 | 2.33 | 1.95 | ||||||||||||||
| V 26018.2 | 2.11? | 1.62 | 2.52 | 1.71 | ||||||||||||||||
| V 26019.1 | 2.27 | 2.09 | 1.08 | |||||||||||||||||
| V 26019.2 | 1.75 | 2.54 | 1.60 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||
| N | 4 | 3 | 3 | 4 | 4 | 4 | ||||||||||||||
| Max | 2.27 | 2.13 | 1.08 | 1.93 | 2.87 | 2.05 | ||||||||||||||
| Min | 1.90 | 1.86 | 1.03 | 1.62 | 2.33 | 1.60 | ||||||||||||||
| Mean | 2.14 | 2.03 | 1.06 | 1.73 | 2.57 | 1.83 | ||||||||||||||
| OrdolagusMCD_2 | V 6268.1 | 2.50 | 2.70 | 0.93 | 1.93 | 3.20 | 2.40 | 2.43 | 3.20 | 2.23 | 2.63 | 3.40 | 2.88 | |||||||
| V 6268.2 | 2.82 | 2.62 | 1.08 | 2.21 | 2.74 | 3.56 | 2.51 | |||||||||||||
| V 6268.6 | 2.39? | |||||||||||||||||||
| V 26016 | 2.36 | 2.64 | 0.89 | 1.90 | 2.90 | 2.14 | 2.50 | 3.14 | 2.24 | |||||||||||
| V 26020 | 2.36 | 2.30? | 1.03? | 2.26 | 3.19 | 2.14 | ||||||||||||||
| V 26021.1 | 2.57 | 2.51 | 1.02 | |||||||||||||||||
| V 26021.2 | 2.38 | 2.43 | 0.98 | |||||||||||||||||
| V 26021.3 | 1.86 | |||||||||||||||||||
| V 26021.4 | 2.38 | 3.26 | 2.19 | |||||||||||||||||
| V 26022.1 | 2.50 | 2.35 | 1.06 | 2.15 | 3.45 | 2.48 | 2.61 | 3.85 | 2.50 | |||||||||||
| V 26023.1 | 2.59 | 2.49 | 1.04 | |||||||||||||||||
| V 26023.2 | 2.56 | 3.25 | 2.54 | |||||||||||||||||
| V 26023.3 | 2.72 | |||||||||||||||||||
| V 26024.1 | 2.19 | 2.17 | 1.01 | |||||||||||||||||
| N | 10 | 9 | 9 | 5 | 3 | 3 | 6 | 6 | 6 | 3 | 2 | 2 | ||||||||
| Max | 2.82 | 2.70 | 1.08 | 2.21 | 3.45 | 2.48 | 2.74 | 3.85 | 2.54 | 2.72 | 3.40 | 2.88 | ||||||||
| Min | 2.19 | 2.17 | 0.89 | 1.86 | 2.90 | 2.14 | 2.26 | 3.14 | 2.14 | 2.38 | 3.26 | 2.19 | ||||||||
| Mean | 2.47 | 2.49 | 1.00 | 2.01 | 3.18 | 2.34 | 2.52 | 3.37 | 2.36 | 2.57 | 3.33 | 2.53 | ||||||||
Table 1 Teeth measurements of Ordolagus from Nei Mongol, China (mm)
| p3 | p4 | m1 | m2 | m3 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | W | L/W | L | W | Wtal | L | W | Wtal | L | W | Wtal | L | W | Wtal | ||||||
| Ordolagus teilhardi | AMNH 20236 | 2.05 | 2.27 | 0.90 | 2.08 | 2.77 | 2.02 | 2.37 | 2.91 | 2.27 | ||||||||||
| IVPP V 26012.1 | 1.93 | 2.35 | 0.82 | 1.93 | 2.60 | 2.05 | 2.25 | 2.80 | 2.00 | 2.50 | 2.88 | 2.33 | 1.58 | 1.90 | 1.05 | |||||
| V 26012.2 | 1.87 | 2.36 | 0.79 | 2.05 | 2.72 | 2.00 | ||||||||||||||
| V 26013.1 | 1.90 | 2.33 | 0.82 | 2.03 | 2.68 | 1.93 | 2.55 | 3.18 | 2.15 | 2.75 | 3.10 | 2.50 | ||||||||
| V 26013.2 | 2.05 | 2.29 | 0.90 | 2.05 | 2.43 | 1.76 | 2.55 | 2.91 | 2.12 | 1.10 | 1.62 | 0.83 | ||||||||
| V 26014 | 2.16 | 2.40 | 0.90 | 1.85 | 2.70 | 1.90 | 2.23 | 3.06 | 1.91 | 2.20 | 3.12 | |||||||||
| V 26015.1 | 1.88 | 2.20 | 0.85 | |||||||||||||||||
| V 26015.2 | 1.80 | 2.75 | 2.05 | |||||||||||||||||
| V 26015.3 | 2.20 | 2.85 | 2.13 | |||||||||||||||||
| V 26017 | 1.57 | 1.71 | 0.92 | |||||||||||||||||
| V 26017* | 2.02* | 2.31* | 0.88 | |||||||||||||||||
| V 26075.1 | 1.93 | 2.46 | 0.78 | 1.98 | 2.73 | 2.22 | ||||||||||||||
| V 26075.3 | 1.88* | |||||||||||||||||||
| V 26075.4 | 2.10* | |||||||||||||||||||
| V 26075.5 | 1.76* | |||||||||||||||||||
| N | 13 | 10 | 10 | 8 | 8 | 8 | 5 | 5 | 5 | 4 | 4 | 3 | 2 | 2 | 2 | |||||
| Max | 2.16 | 2.46 | 0.92 | 2.08 | 2.77 | 2.22 | 2.55 | 3.18 | 2.27 | 2.75 | 3.12 | 2.50 | 1.58 | 1.90 | 1.05 | |||||
| Min | 1.57 | 1.71 | 0.78 | 1.80 | 2.43 | 1.76 | 2.20 | 2.80 | 1.91 | 2.20 | 2.88 | 2.12 | 1.10 | 1.62 | 0.83 | |||||
| Mean | 1.93 | 2.27 | 0.86 | 1.97 | 2.67 | 1.99 | 2.32 | 2.96 | 2.09 | 2.50 | 3.00 | 2.31 | 1.34 | 1.76 | 0.94 | |||||
| OrdolagusMCD_1 | V 6268.13 | 2.25 | 2.13 | 1.06 | 1.93 | 2.87 | 2.05 | 2.23 | 3.17 | 1.98 | 2.52 | 3.22 | 2.30 | |||||||
| V 26018.1 | 1.90 | 1.86 | 1.03 | 1.62 | 2.33 | 1.95 | ||||||||||||||
| V 26018.2 | 2.11? | 1.62 | 2.52 | 1.71 | ||||||||||||||||
| V 26019.1 | 2.27 | 2.09 | 1.08 | |||||||||||||||||
| V 26019.2 | 1.75 | 2.54 | 1.60 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||
| N | 4 | 3 | 3 | 4 | 4 | 4 | ||||||||||||||
| Max | 2.27 | 2.13 | 1.08 | 1.93 | 2.87 | 2.05 | ||||||||||||||
| Min | 1.90 | 1.86 | 1.03 | 1.62 | 2.33 | 1.60 | ||||||||||||||
| Mean | 2.14 | 2.03 | 1.06 | 1.73 | 2.57 | 1.83 | ||||||||||||||
| OrdolagusMCD_2 | V 6268.1 | 2.50 | 2.70 | 0.93 | 1.93 | 3.20 | 2.40 | 2.43 | 3.20 | 2.23 | 2.63 | 3.40 | 2.88 | |||||||
| V 6268.2 | 2.82 | 2.62 | 1.08 | 2.21 | 2.74 | 3.56 | 2.51 | |||||||||||||
| V 6268.6 | 2.39? | |||||||||||||||||||
| V 26016 | 2.36 | 2.64 | 0.89 | 1.90 | 2.90 | 2.14 | 2.50 | 3.14 | 2.24 | |||||||||||
| V 26020 | 2.36 | 2.30? | 1.03? | 2.26 | 3.19 | 2.14 | ||||||||||||||
| V 26021.1 | 2.57 | 2.51 | 1.02 | |||||||||||||||||
| V 26021.2 | 2.38 | 2.43 | 0.98 | |||||||||||||||||
| V 26021.3 | 1.86 | |||||||||||||||||||
| V 26021.4 | 2.38 | 3.26 | 2.19 | |||||||||||||||||
| V 26022.1 | 2.50 | 2.35 | 1.06 | 2.15 | 3.45 | 2.48 | 2.61 | 3.85 | 2.50 | |||||||||||
| V 26023.1 | 2.59 | 2.49 | 1.04 | |||||||||||||||||
| V 26023.2 | 2.56 | 3.25 | 2.54 | |||||||||||||||||
| V 26023.3 | 2.72 | |||||||||||||||||||
| V 26024.1 | 2.19 | 2.17 | 1.01 | |||||||||||||||||
| N | 10 | 9 | 9 | 5 | 3 | 3 | 6 | 6 | 6 | 3 | 2 | 2 | ||||||||
| Max | 2.82 | 2.70 | 1.08 | 2.21 | 3.45 | 2.48 | 2.74 | 3.85 | 2.54 | 2.72 | 3.40 | 2.88 | ||||||||
| Min | 2.19 | 2.17 | 0.89 | 1.86 | 2.90 | 2.14 | 2.26 | 3.14 | 2.14 | 2.38 | 3.26 | 2.19 | ||||||||
| Mean | 2.47 | 2.49 | 1.00 | 2.01 | 3.18 | 2.34 | 2.52 | 3.37 | 2.36 | 2.57 | 3.33 | 2.53 | ||||||||
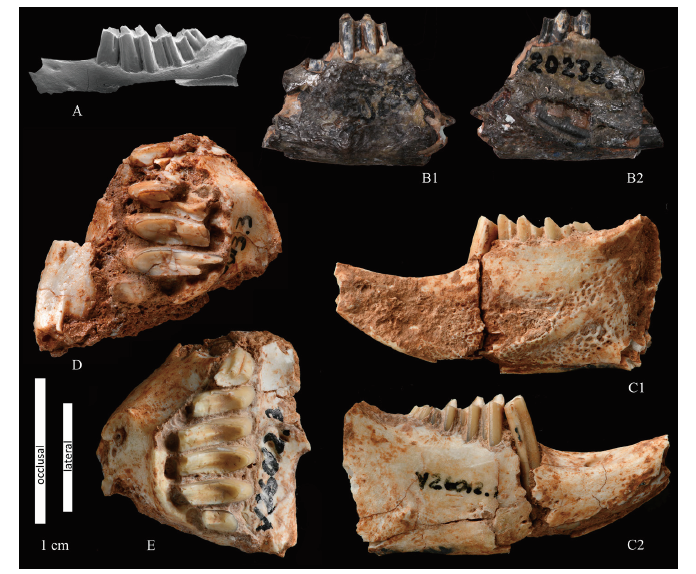
Fig. 3 Gobiolagus tolmachovi and Ordolagus teilhardi from Nei Mongol, China A. Gobiolagus tolmachovi: left lower jaw in labial view, AMNH 26142, Nei Mongol, late Middle Eocene (Burke, 1941; Meng et al., 2005); B-E. Ordolagus teilhardi: B. left lower jaw in labial (B1) and lingual (B2) views, holotype, AMNH 20236; C. right lower jaw in lingual (C1) and labial (C2) views, IVPP V 26012.1; D. left upper jaw in occlusal view, V 26013.3; E. right upper jaw in occlusal view, V 26012.3

Fig. 4 Camera lucida drawings of teeth of Ordolagus from Nei Mongol, China A-J. Ordolagus teilhardi: A. right p3-m3, IVPP V 26012.1; B. left p3-p4, V 26012.2; C. left p3-m1, V 26015.1-3; D. left p3-m1, V 26013.1; E. right p3-p4+m2-m3, V 26013.2; F. right p3-m2, V 26014; G1. right p3-m2, V 26017; G2. detail of p3 in lateral lingual view, V 26017; G3. detail of m2 in lateral lingual view, V 26017; H. left p3, V 26075.2; I. right p3, V 26075.3; J. detail of p3-p4 of left lower jaw, V 26075.1; K-M. Ordolagus MDC_1: K. left p3-m2, V 6268.13; L1. left p3-p4, V 26019.1, V 26019.2; L2. left p3 in lateral labial view, V 26019.1; M. right p3-p4, V 26018.1; N-V. Ordolagus MDC_2: N. right dp3, V 26022.2; O. left dp4, V 26022.3; P. right p3, V 26021.1; Q1. right p3-m2, V 6268.1; Q2. detail of p3 in lateral labial view, V 6268.1; R. left p3, V 26023.1; S. right p3, V 26024.1; T. right p3, V 26021.2; U. left p3, V 26020; V. left p3-m1, V 26022.1. Occlusal surfaces drawn parallel to the observer
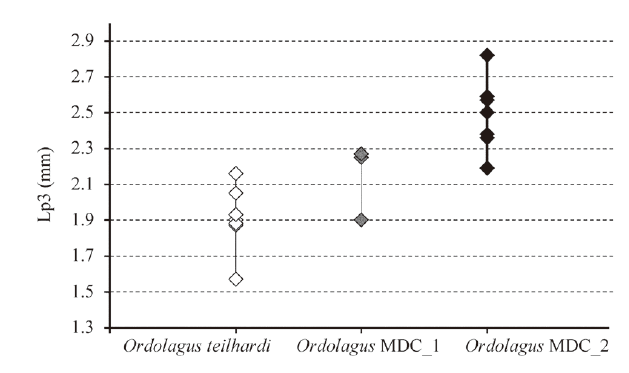
Fig. 5 Length of p3 (in mm) of Ordolagus teilhardi (white diamonds), Ordolagus MDC_1 (grey diamonds) and Ordolagus MDC_2 (black diamonds)
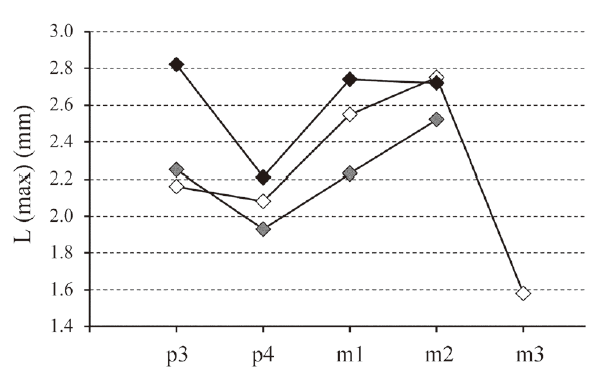
Fig. 6 Length of the lower teeth (in mm) in Ordolagus teilhardi (white diamonds), Ordolagus MDC_1 (grey diamonds) and Ordolagus MDC_2 (black diamonds) The considered values are the maximum L values for each tooth position
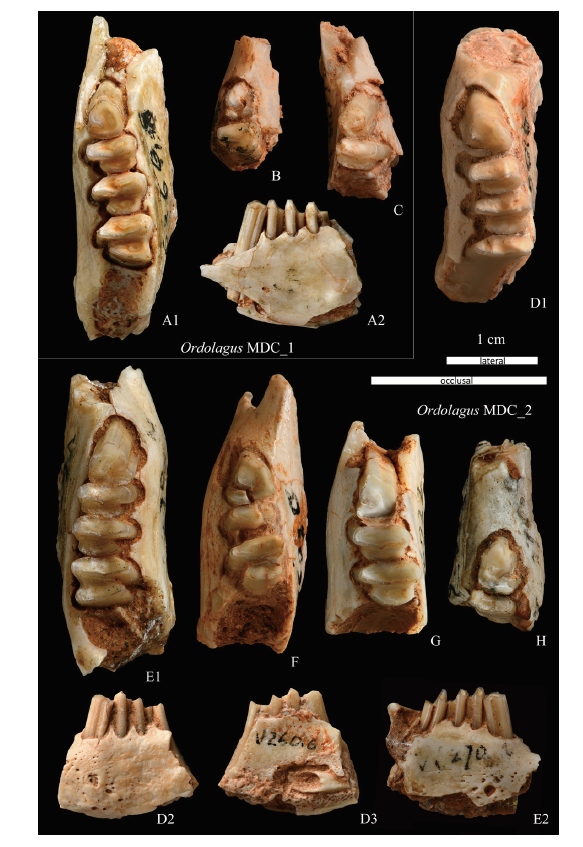
Fig. 7 Ordolagus MDC_1 and Ordolagus MDC_2 from Nei Mongol, China A-C. Ordolagus MDC_1: A. left lower jaw in occlusal (A1) and labial (A2) views, IVPP V 6268.13; B. left lower jaw, V 26018.2; C. right lower jaw, V 26018.1. D-H. Ordolagus MDC_2: D. left lower jaw in occlusal (D1), labial (D2), and lingual (D3) views, V 26016; E. right lower jaw in occlusal (E1) and labial (E2) views, V 6268.1; F. left lower jaw, V 26020; G. left lower jaw, V 26022.1; H. right lower jaw, V 6268.6
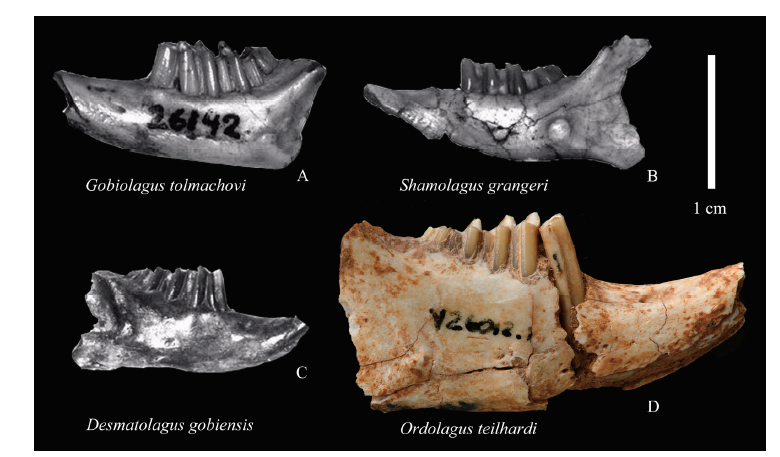
Fig. 8 Size comparison between lower jaws of Ordolagus and other selected Eocene-Early Oligocene Asian lagomorphs A. Gobiolagus tolmachovi, AMNH 26142, left lower jaw with p3-m3, from Meng et al., 2005:4, fig. 1.1; B. Shamolagus grangeri, AMNH 26289, left lower jaw with p4-m3, from Meng et al., 2005:4, fig. 1.6; C. Desmatolagus gobiensis, AMNH 19102, right lower jaw with p3-m3, from Meng et al., 2005:4, fig 1.9; D. Ordolagus teilhardi, IVPP V 26012.1, right lower jaw with p3-m3
| [1] | Angelone C, 2005. Evolutionary trends of the genus Prolagus (Ochotonidae, Lagomorpha) in the Mediterranean islands. In: Alcover J A, Bover P eds. Proceedings of the International Symposium “Insular Vertebrate Evolution: the Palaeontological Approach”. Mon Soc Hist Nat Balears, 12:17-26 |
| [2] |
Angelone C, 2009a. Lagomorphs from the Middle Miocene of Sandelzhausen (southern Germany). Paläont Z, 83:67-75
DOI URL |
| [3] | Angelone C, 2009b. The early Vallesian vertebrates of Atzelsdorf (Late Miocene, Austria) - 6. Lagomorpha. Ann Naturhist Mus Wien, Ser A, 111:515-518 |
| [4] | Angelone C, Schultz J A, Erbajeva M A, 2014. Determining the ontogenetic variation of lower cheek teeth occlusal surface patterns in lagomorphs using Micro CT-technology - preliminary results and perspectives. Paleont Electron, 17.1.5 |
| [5] |
Angelone C, Čermák S, Moncunill-Solé B et al., 2018. Systematics and paleobiogeography of Sardolagus obscurus n. gen. n. sp. (Leporidae, Lagomorpha) from the Early Pleistocene of Sardinia. J Paleont, 92:506-522
DOI URL |
| [6] | Ao H, Dupont-Nivet G, Rohling E et al., 2020. Orbital climate variability on the northeastern Tibetan Plateau across the Eocene-Oligocene transition. Nat Commun, 11: 10.1038/s41467-020-18824-8 |
| [7] |
Bai B, Wang Y Q, Zhang Z Q, 2018. The Late Eocene hyracodontid perissodactyl Ardynia from Saint Jacques, Inner Mongolia, China and its implications for the potential Eocene-Oligocene boundary. Palaeoworld, 27(2):247-257
DOI URL |
| [8] | Bohlin B, 1942. The fossil mammals from the Tertiary deposit of Taben-buluk, western Kansu. Part I. Insectivora and Lagomorpha. Palaeont Sin New Ser C, 8A:1-113 |
| [9] | Brandt J F, 1855. Beiträge zur näheren Kenntniss der Säugethiere Russlands. Mem Acad Sci St Petersb, Phys, Mat, Nat, 6(9):1-365 |
| [10] | Bucher H, 1982. Étude des genres Marcuinomys Lavocat et Lagopsis Schlosser (Lagomorpha, Mammalia) du Miocène inférieur et moyen de France. Implications biostratigraphiques et phylogénétiques. Bull Mus Hist Nat Paris, 4:43-74 |
| [11] | Burke J J, 1941. New fossil Leporidae from Mongolia. Am Mus Novit, 1117:1-23 |
| [12] | Dawson M R, 1958. Later Tertiary Leporidae of North America. Univ Kansas Paleont Contrib Vertebr, 6:1-75 |
| [13] | Dawson M R, 1967. Lagomorph history and the stratigraphic record. Raymond C. Moore commemorative volume. Univ Kansas Dept Geol Spec Publ, 2:287-316 |
| [14] | Dawson M R, 2008. 17. Lagomorpha. In: Janis C M, Gunnell G F, Uhen M D eds. Evolution of Tertiary Mammals of North America, Vol.2,Small Mammals,Xenarthrans, and Marine Mammals. Cambridge: Cambridge University Press. 293-310 |
| [15] | Daxner-Höck G, Badamgarav D, Barsbold R et al., 2017. Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia). Paleobiodiv Paleoenv, 97:111-218 |
| [16] | De Muizon C, 1977. Revision des lagomorphes des couches à Baluchitherium (Oligocène supérieur) de San-tao-ho (Ordos, Chine). Bull Mus Natl Hist Nat, Sci Terre, 65:265-294 |
| [17] |
Emry R J, Gawne C E, 1986. A primitive, Early Oligocene species of Palaeolagus (Mammalia, Lagomorpha) from the Flagstaff Rim area of Wyoming. J Vert Paleont, 6(3):271-280
DOI URL |
| [18] | Erbajeva M A, 2007. Oligocene-Miocene vertebrates from the Valley of Lakes (Central Mongolia): morphology, phylogenetic and stratigraphic implications. 5. Lagomorpha (Mammalia): preliminary results. Ann Naturhist Mus Wien Ser A, 108:165-171 |
| [19] | Erbajeva M A, 2012. Biodiversity and evolutionary development of Oligocene-Pliocene lagomorphs (Lagomorpha, Mammalia) of Mongolia. Erforsch Biol Ress Mongol, 12:47-54 |
| [20] | Erbajeva M A, Daxner-Höck G, 2001. Paleogene and Neogene lagomorphs from the Valley of Lakes, central Mongolia. Lynx New Ser, 32:55-65 |
| [21] | Erbajeva M A, Daxner-Höck G, 2014. The most prominent Lagomorpha from the Oligocene and Early Miocene of Mongolia. Ann Naturhist Mus Wien Ser A, 116:215-245 |
| [22] | Erbajeva M A, Şen Ş, 1988. Systematic of some Oligocene Lagomorpha (Mammalia) from China. Neues Jahrb Geol Paläont Monatsh, (2):95-105 |
| [23] | Fischer de Waldheim J G, 1817. Adversaria zoological. Mém Soc Imp Nat Moscow, 5:368-428 |
| [24] | Fortelius M, Eronen J, Jernval J et al., 2002. Fossil mammals resolve regional patterns of Eurasian climate change over 20 million years. Evol Ecol Res, 4:1005-1016 |
| [25] | Fostowicz-Frelik Ł, 2013. Reassessment of Chadrolagus and Litolagus (Mammalia, Lagomorpha) and a new genus of North American Eocene lagomorph from Wyoming. Am Mus Novit, 3773:1-76 |
| [26] | Fostowicz-Frelik Ł, Li Q, 2014. A new genus of stem lagomorph (Mammalia: Glires) from the Middle Eocene of the Erlian Basin, Nei Mongol, China. Acta Zool Cracov, 57:29-42 |
| [27] |
Fostowicz-Frelick Ł, Tabrum A R, 2009. Leporids (Mammalia, Lagomorpha) from the Diamond O Ranch Local Fauna, latest Middle Eocene of southwestern Montana. Ann Carnegie Mus, 78(3):253-271
DOI URL |
| [28] | Fostowicz-Frelik Ł, Li C K, Meng J et al., 2012. New Gobiolagus (Mammalia: Lagomorpha) material from the Middle Eocene of Erden Obo (Nei Mongol, China). Vert PalAsiat, 50:219-236 |
| [29] | Fostowicz-Frelik Ł, Li C K, Li Q et al., 2015. Strenulagus (Mammalia: Lagomorpha) from the Middle Eocene Irdin Manha Formation of the Erlian Basin, Nei Mongol, China. Acta Geol Sin, 89:12-26 |
| [30] | Gawne C E, 1978. Leporids (Lagomorpha, Mammalia) from the Chadronian (Oligocene) deposits of Flagstaff Rim, Wyoming. J Vert Paleont, 52(5):1103-1118 |
| [31] |
Ge D, Wen Z, Xia L et al., 2013. Evolutionary history of lagomorphs in response to global environmental change. PLoS ONE, 8(4):e59668
DOI URL |
| [32] |
Green M, 1942. A study of the Oligocene Leporidae in the Kansas University Museum of Vertebrate Paleontology. Trans Kansas Acad Sci, 45:229-247
DOI URL |
| [33] | Gureev A A, 1964. Fauna of USSR, Mammals, Lagomorphs. Vol. 3(10). Moscow-Leningrad: Nauka Press. 1-276 |
| [34] |
Harzhauser M, Daxner-Höck G, López-Guerrero P et al., 2016. Stepwise onset of the Icehouse world and its impact on Oligocene-Miocene Central Asian mammals. Sci Rep, 6:36169
DOI URL |
| [35] | Hauptvogel D W, Pekar S F, Pincay V, 2017. Evidence for a heavily glaciated Anctartica during the Late Oligocene “warming” (27.8-24.5 Ma): stable ispotope records from ODP site 690. Paleoceanography, 32, doi: 10/1002/2016PA002972 |
| [36] |
Head M J, Aubry M P, Walker M et al., 2017. A case for formalizing subseries (subepochs) of the Cenozoic. Episodes, 40:22-27
DOI URL |
| [37] |
Hooker J J, Collinson M E, Sille N P, 2004. Eocene-Oligocene mammalian faunal turnover in the Hampshire Basin, UK: calibration to the global time scale and the major cooling event. J Geol Soc London, 161:161-172
DOI URL |
| [38] | Huang X S, 1982. Preliminary observations on the Oligocene deposits and mammalian fauna from Alashan Zuoqi, Nei Mongol. Vert PalAsiat, 20:337-349 |
| [39] | Huang X S, 1986. Fossil leporids from the Middle Oligocene of Ulantatal, Nei Mongol. Vert PalAsiat, 24:274-284 |
| [40] |
Kohn M J, Josef J A, Madden R et al., 2004. Climate stability across the Eocene-Oligocene transition, southern Argentina. Geology, 32:621-624
DOI URL |
| [41] |
Kohn M J, Strömberg C A E, Madden R H et al., 2015. Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeogr Palaeoclimatol Palaeoecol, 435:24-37
DOI URL |
| [42] |
Kraatz P, Geisler J H, 2010. Eocene-Oligocene transition in Central Asia and its effects on mammalian evolution. Geology, 38:111-114
DOI URL |
| [43] | Li C K, 1965. Eocene leporids of North China. Vert PalAsiat, 9:23-33 |
| [44] | Li C K, Zhang Z Q, 2019. Glires I:Duplicidentata, Simplicidentata-Mixodontia. Palaeovertebrata Sinica, 3(4). Beijing: Science Press. 1-217 |
| [45] |
Lopatin A V, Averianov A O, 2006. Eocene Lagomorpha (Mammalia) of Asia. 2. Strenulagus and Gobiolagus (Strenulagidae). Paleontol J, 40(2):198-206
DOI URL |
| [46] | López Martínez N, 1974. Évolution de la lignée d’Ochotonidés Piezodus-Prolagus dans le Cénozoïque d’Europe Sud-Occidentale. Ph.D Thesis. Montpellier: Université des Sciences et Techniques du Languedoc. 1-165 |
| [47] | López Martínez N, 1989. Revisión sistemática y biostratigráfica de los Lagomorpha (Mammalia) del Terciario y Cuaternario de España. Mem Mus Paleont Univ Zaragoza, 3:1-350 |
| [48] | López Martínez N, 2008. The lagomorph fossil record and the origin of the European rabbit. In: Alves P C, Ferrand N, Hackländer K eds. Lagomorph Biology. Evolution, Ecology, Conservation. Berlin-Heidelberg: Springer. 27-46 |
| [49] | López Martínez N, Thaler L, 1975. Biogéographie, évolution et complements à la systématique du groupe d’Ochotonidés Piezodus-Prolagus (Mammalia, Lagomorpha). Bull Soc Géol Fr, 17:850-866 |
| [50] | López Martínez N, Likius A, Mackaye H T et al., 2007. A new lagomorph from the Late Miocene of Chad (Central Africa). Rev Esp Paleont, 22:1-20 |
| [51] | Lyon M W, 1904. Classification of the hares and their allies. Smithson Miscell Coll, 45:321-463 |
| [52] | Major C J F, 1899. On fossil and recent Lagomorpha. Trans Linn Soc London, 2(7):433-520 |
| [53] | Mein P, Adrover R, 1982. Une faunule de mammifères insulaires dans le Miocène moyen de Majorque (Îles Baléares). Geobios, Mém Spec, 6:451-463 |
| [54] | Mellet J S, 1968. The Oligocene Hsanda Gol Formation, Mongolia: a revised faunal list. Am Mus Novit, 2318:1-16 |
| [55] |
Meng J, McKenna M, 1998. Faunal turnovers of Paleogene mammals from the Mongolian Plateau. Nature, 394:364-367
DOI URL |
| [56] | Meng J, Hu Y M, Li C K, 2005. Gobiolagus (Lagomorpha, Mammalia) from Eocene Ula Usu, Inner Mongolia, and comments on Eocene lagomorphs of Asia. Palaeont Electron, 88(1):7A |
| [57] | Moncunill-Solé B, Tuveri C, Arca M et al., 2016. Comparing the body mass variations in endemic insular species of the genus Prolagus (Ochotonidae, Lagomorpha) in the Pleistocene of Sardinia (Italy). Riv Ital Paleont Stratigr, 122:25-36 |
| [58] |
Pound M J, Salzmann U, 2017. Heterogeneity in global vegetation and terrestrial climate change during the Late Eocene to Early Oligocene transition. Sci Rep, 7:43386
DOI URL |
| [59] |
Prothero D, Heaton T, 1996. Faunal stability during the Early Oligocene climatic crash. Palaeogeogr Palaeoclimatol Palaeoecol, 127:257-283
DOI URL |
| [60] | Qiu Z D, Wang X M, Li Q, 2006. Faunal succession and biochronology of the Miocene through the Pliocene in Nei Mongol (Inner Mongolia). Vert PalAsiat, 44(2):164-181 |
| [61] |
Richoz S, Baldermann A, Frauwallner A et al., 2017. Geochemistry and mineralogy of the Oligo-Miocene sediments of the Valley of Lakes, Mongolia. Palaeobiodiv Palaeoenv, 97:233-258
DOI URL |
| [62] | Şen Ş, 2003. Lagomorpha. In: Fortelius M, Kappelman J, Şen Ş et al. eds. Geology and Paleontology of the Miocene Sinap Formation, Turkey. New York: Columbia University Press. 163-178 |
| [63] | Stehlin H G, 1910. Remarques sur les faunules de Mammifères des couches éocènes et oligocènes du Bassin de Paris. Bull Soc Géol Fr, 9:488-520 |
| [64] | Sun J, Windley B F, 2015. Onset of aridification by 34 Ma across the Eocene-Oligocene transition in Central Asia. Geology, 43(11): G37165.1 |
| [65] | Tabrum A R, Fostowicz-Frelik Ł, 2008. Lagomorphs from the late Duchesnean (late Middle Eocene) Diamond O Ranch local fauna of southwestern Montana. J Vert Paleont, 28(Suppl 3):150A |
| [66] | Teilhard de Chardin P, 1926. Description des mammifères tertiaires de Chine et de Mongolie. Ann Paléont, 15:1-52 |
| [67] | Tobien H, 1974. Zur Gebißstruktur, Systematik und Evolution der Genera Amphilagus und Titanomys (Lagomorpha, Mammalia) aus einigen Vorkommen im jüngeren Tertiär Mittelund Westeuropas. Mainzer Geowiss Mitt, 3:95-214 |
| [68] | Tobien H, 1975. Zur Gebißstruktur, Systematik und Evolution der Genera Piezodus, Prolagus und Ptychoprolagus (Lagomorpha, Mammalia), aus einigen Vorkommen im jüngeren Tertiär Mittel- und Westeuropas. Notiz Hess Landesamt Bodenforsch Wiesbaden, 103:103-186 |
| [69] | Tobien H, 1978. Brachydonty and hypsodonty in some Paleogene Eurasian lagomorphs. Mainzer Geowiss Mitt, 6:161-175 |
| [70] | Tong Y S, Lei Y Z, 1987. Fossil lagomorphs (Mammalia) from the Hetaoyuan Eocene of Xichuan, Hennan. Vert PalAsiat, 25:208-221 |
| [71] | Vianey-Liaud M, Schmidt-Kittler N, Marivaux L, 2006. The Ctenodactylidae (Rodentia) from the Oligocene of Ulantatal (Inner Mongolia, China). Paleovertebrata, 34:111-206 |
| [72] | Wang B Y, 1997. Chronological sequence and subdivision of Chinese Oligocene mammalian faunas. J Stratigr, 21:183-191 |
| [73] | Wang J, Zhang Z Q, 2015. Phylogenetic analysis on Palaeogale (Palaeolagidae, Carnivora) based on specimens from Oligocene strata of Saint-Jacques, Nei Mongol. Vert PalAsiat, 53(4):310-334 |
| [74] | Wasiljeff J, Kaakinen A, Salminen J et al., 2020. Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: implications for Asian aridification and faunal turnover before the Eocene-Oligocene boundary. Earth Planet Sci Lett, 535:116-125 |
| [75] |
Wood A E, 1940. The mammalian fauna of the White River Oligocene. Part III: Lagomorpha. Trans Am Philos Soc, 28:271-362
DOI URL |
| [76] |
Zachos J, Pagani M, Sloan L et al., 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Nature, 292:686-693
DOI URL |
| [77] | Zhang Z Q, Dawson M R, Huang X S, 2001. A new species of Gobiolagus (Lagomorpha, Mammalia) from the Middle Eocene of the Shanxi Province, China. Ann Carnegie Mus, 70:257-261 |
| [78] |
Zhang Z Q, Liu Y, Wang L H et al., 2016. Lithostratigraphic context of Oligocene mammalian faunas from Ulantatal, Nei Mongol, China. C R Palevol, 15(7):903-910
DOI URL |
| [1] | 李琪, 李茜. 内蒙古二连盆地沙拉木伦期啮齿类动物群研究. 古脊椎动物学报, 2023, 61(1): 43-70. |
| [2] | Joonas Wasiljeff, 张兆群. 内蒙古阿拉善左旗乌兰塔塔尔最晚始新世-渐新世剖面的区域年代地层意义. 古脊椎动物学报, 2022, 60(1): 42-53. |
| [3] | 王世骐, 李春晓, 张晓晓. 乳齿象类的命名与中文译法及分类学问题讨论. 古脊椎动物学报, 2021, 59(4): 295-332. |
| [4] | 王倩, 刘艳, 王李花, 傅明楷, 张兆群. 内蒙古韩家营玄武岩夹层中三趾马动物群化石. 古脊椎动物学报, 2021, 59(2): 125-137. |
| [5] | 张立民, 董为, 倪喜军, 李强. 晚中新世晚期土城子小哺乳动物组合及土城子动物群在内蒙古中部地区新近纪哺乳动物群序列中的位置. 古脊椎动物学报, 2021, 59(1): 45-63. |
| [6] | 邱铸鼎, 王晓鸣, 李强, 李录, 王洪江, 陈海峰. 内蒙古哈拉津胡舒晚中新世动物群. 古脊椎动物学报, 2021, 59(1): 19-26. |
| [7] | 李茜. 内蒙古二连盆地额尔登敖包剖面新增查干鼠类、圆柱齿鼠类及梳趾鼠类化石材料. 古脊椎动物学报, 2021, 59(1): 1-18. |
| [8] | 刘玉海, 朱敏, 林翔鸿, 卢立伍, 盖志琨. 新疆塔里木盆地志留纪盔甲鱼类新知. 古脊椎动物学报, 2019, 57(4): 253-273. |
| [9] | Lawrence J. FLYNN, 金昌柱, Jay KELLEY, Nina G. JABLONSKI, 吉学平, Denise F. SU, 邓涛, 李强. 云南晚中新世苏门答腊兔(Nesolagus)分离时间的标定. 古脊椎动物学报, 2019, 57(3): 214-224. |
| [10] | 刘俊. 二叠纪脑包沟组的四足动物群——4. 二齿兽类的多样性. 古脊椎动物学报, 2019, 57(3): 173-180. |
| [11] | 白 滨, 王元青, 孟 津. 内蒙古二连盆地沙拉木伦地区中始新世全脊貘(Teleolophus)(奇蹄目:貘超科)头后骨骼研究. 古脊椎动物学报, 2018, 56(3): 193-215. |
| [12] | 董 为, 刘文晖, 张立民, 白炜鹏, 蔡保全. 内蒙古化德土城子地点晚中新世麝科化石新材料. 古脊椎动物学报, 2018, 56(3): 229-247. |
| [13] | 张颖奇,金昌柱,王 元,Alejandra ORTIZ,何 锴,Terry HARRISON. 广西崇左更新世长臂猿化石新材料. 古脊椎动物学报, 2018, 56(3): 248-263. |
| [14] | 董 为,刘文晖,张立民,蔡保全. 内蒙古化德土城子地点晚中新世鹿科化石新材料. 古脊椎动物学报, 2018, 56(2): 157-175. |
| [15] | Thomas A. STIDHAM,王元青 . 内蒙古中始新世一似ameghinornithid鸟类(鸟纲: Cariamae: Ameghinornithidae?). 古脊椎动物学报, 2017, 55(3): 218-226. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||