
古脊椎动物学报 ›› 2019, Vol. 57 ›› Issue (1): 1-37.DOI: 10.19615/j.cnki.1000-3118.180530
• • 下一篇
王敏1,2( ), 邹晶梅1,2, 周忠和1,2
), 邹晶梅1,2, 周忠和1,2
收稿日期:2018-02-05
出版日期:2019-01-20
发布日期:2019-01-20
基金资助:
WANG Min1,2(), Jingmai O’CONNOR1,2, ZHOU Zhong-He1,2
Received:2018-02-05
Published:2019-01-20
Online:2019-01-20
Contact:
wangmin@ivpp.ac.cn摘要:
孔子鸟目是早白垩世鸟类中具有角质喙和尾综骨的最原始的鸟类类群之一。在过去近20年的研究历史中,千余件孔子鸟类标本被发现,其数目多过其他中生代鸟类和非鸟类恐龙。目前,共计4个属,10个种被先后归入到孔子鸟目中,然而这些属种的分类位置是否有效尚不清楚。通过对已发表的所有孔子鸟类属种进行详细的对比研究,证实了其中多数属种的鉴定特征有误,这些特征或受保存的影响,或代表了种内和个体发育的变化,亦或广泛分布在孔子鸟类或基干鸟类中,而并不能成为这些属种建属或建种的依据。认为此前命名的孙氏孔子鸟、费氏孔子鸟、义县锦州鸟、张吉营锦州鸟和建昌孔子鸟均属于圣贤孔子鸟的同物异名;而川州孔子鸟未保存确切的鉴定特征而难以和其他孔子鸟类相区别,暂将其归入孔子鸟科未定种。至此,对所有已报道的孔子鸟目的分类厘定认为,目前孔子鸟目共包括1科、3属、4种,分别是圣贤孔子鸟、杜氏孔子鸟、横道子长城鸟和郑氏始孔子鸟,并对这些属种各自的鉴定特征进行了修正。
中图分类号:
王敏, 邹晶梅, 周忠和. 孔子鸟目(鸟类:尾综骨类)的分类厘定. 古脊椎动物学报, 2019, 57(1): 1-37.
WANG Min, Jingmai O’CONNOR, ZHOU Zhong-He. A taxonomical revision of the Confuciusornithiformes (Aves: Pygostylia). Vertebrata Palasiatica, 2019, 57(1): 1-37.

Fig. 1 Skull of Confuciusornis sanctus in comparison with those of C. dui and other Early Cretaceous birds A, C. photograph (A) and line drawing (C) of Confuciusornis sanctus IVPP V 13168 with a separate drawing of the maxilla (right), showing the halberd-shaped dorsal process of the maxilla (considered an autapomorphy here); B. C. sanctus V 13171 with line drawing of the maxilla; D. drawing of C. dui with a separate drawing of the maxilla (modified from Hou et al., 1999); E. Jeholornis sp. V 14978 (Aves, non-Pygostylia); F. Pterygornis dapingfangensis V 16363 (Aves, Enantiornithes). The arrowheads in B denote the rostral notches of the premaxillary and mandibular symphyses. The arrow in B indicates the ventral process of the surangular (considered an autapomorphy here). Scale bars = 10 mm; C, D not to scale

Fig. 2 Detail anatomy of Confuciusornis sanctus showing some diagnostic characters A. right forelimb of C. sanctus IVPP V 10928 (hand in ventral view, humerus in cranial view); B. hindlimbs of C. sanctus V 11552 in dorsal view; C, D. photograph (C) and reconstruction (D) of the sternum of C. sanctus in dorsal view; E. sternal reconstruction of C. dui (modified from Hou et al., 1999); F. sternal reconstruction of Eoconfuciusornis zhengi (modified from Zhang et al., 2008) The arrowhead in A denotes the ridge on the craniaodorsal margin of the deltopectoral crest, and the arrow indicates the unfused carpometacarpus. The arrowheads in B mark the extensor retinaculum tubercle on the dorsal surface of the distal tibiotarsus (diagnostic character); and the arrows indicate the dorsal tubercles on metatarsals II and III. The arrows in D denote the cranial and caudal projections of the lateral process of the sternum in C. sanctus (diagnostic character). The dash lines in E indicate the length (vertical) and width (horizontal) of the sternum that are measured in this study Scale bars equal 10 mm in A and C, 5 mm in B; D-F not to scale
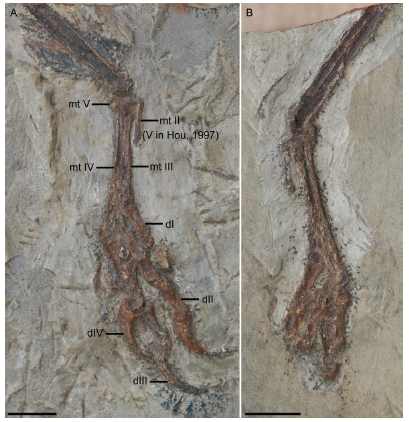
Fig. 3 Photographs of Confuciusornis sanctus [= 'Confuciusornis chuonzhous'], IVPP V 10919 A. slab; B. counter slab. The broken piece of the proximal end of metatarsal II was misinterpreted as metatarsal V by Hou (1997). See text for further explanations. Scale bars = 10 mm
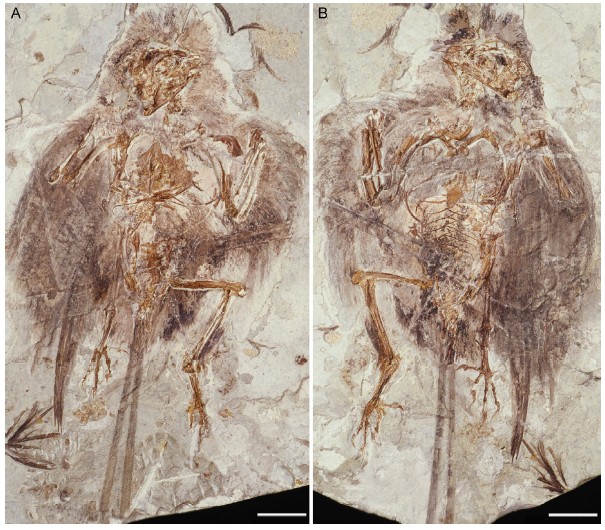
Fig. 4 Photograph of the holotype of Changchengornis hengdaoziensis, GMV 2129 Scale bars = 20 mm
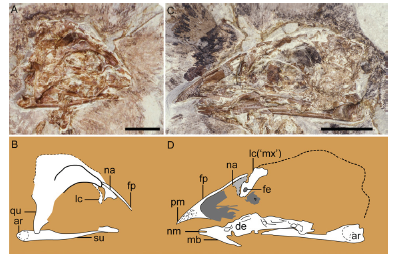
Fig. 5 Cranial morphology of Changchengornis hengdaoziensis, GMV 2129 A, B. skull in right lateral view preserved on GMV 2129a; C, D. skull in left lateral view preserved on GMV 2129b. A, C. photograph; B, D. line drawing. The dorsal process of the maxilla (labeled as 'mx') identified in Chiappe et al. (1999) is reinterpreted to be the lacrimal here (see text for explanation) Scale bars = 10 mm (A, C); line drawings (C, D) are not to scale
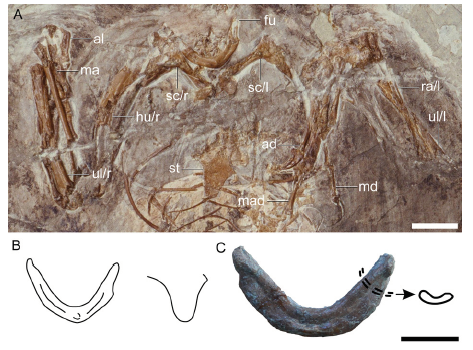
Fig. 6 Pectoral girdle of Changchengornis hengdaoziensis in comparison with Confuciusornis sanctus A. pectoral girdle of Ch. hengdaoziensis in dorsal view; B. line drawing of the furcula (left) and caudal end of the sternum (right) of Ch. hengdaoziensis, showing that the dorsal surface is excavated by a dorsal furrow as in C. sanctus; C. furcula of C. sanctus IVPP V 11521 in dorsal view with drawing of the cross section Scale bars = 10 mm (A, C); line drawings (B, C) are not to scale

Fig. 7 Skeleton and anatomy of Confuciusornis sanctus [= 'Jinzhouornis yixianensis'], IVPP V 14412 A. overview; B. skull in oblique right lateral view; C. proximal left humerus showing the ridge on the cranial surface (arrow), as in C. sanctus; D. lower legs in dorsal view, exhibiting the presences of an extensor retinaculum on distal tibiotarsus (arrowhead), and the proximal ends of metatarsals II and III (arrows), as in C. sanctus. Scale bars = 10 mm

Fig. 8 Skeleton and cranial anatomy of Confuciusornis sanctus [='Jinzhouornis zhangjiyingia'], IVPP V 12352 A. overview; B, C. photograph (B) and line drawing (C) of the skull in left lateral viewThe arrowhead in B denotes the ventral end of the ventral process of the postorbital, which was misinterpreted as the quadratojugal by Hou (1997). Scale bars = 10 mm
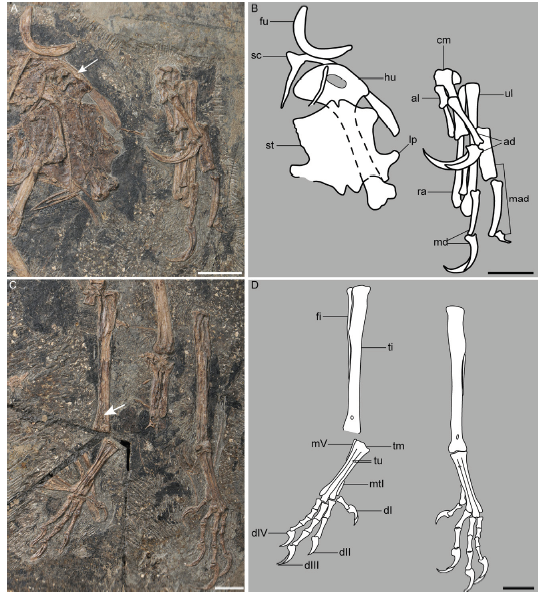
Fig. 9 Detail anatomy of Confuciusornis sanctus [= 'Jinzhouornis zhangjiyingia'], IVPP V 12352 A, B. pectoral girdle, sternum, and left forelimb; C, D. hindlimb showing the presence of an extensor retinaculum on distal tibiotarsus (arrow); A, C. photographs; B, D. line drawing. The arrow in A denotes the ridge on the cranial surface of the deltopectoral crest, as in C. sanctus. Scale bars = 10 mm

Fig. 10 Skeletal anatomy of the holotype of Eoconfuciusornis zhengi, IVPP V 11977 A. overview; B, C. photograph (B) and line drawing (C) of the skull; D. furcula (dorsal view) and proximal part of right humerus (cranial view). The arrowhead in C indicates that the surangular bears a ventral process as in C. sanctus. The arrow in C shows that the dorsal process of the maxilla is craniocaudally compressed Scale bars = 10 mm (A, B, D); line drawing (C) is not to scale
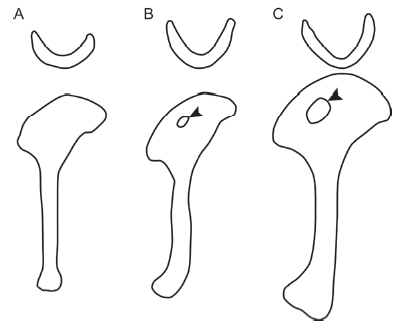
Fig. 11 Comparison of furcula and humerus anatomy between Eoconfuciusornis zhengi and Confuciusornis sanctus A. holotype of Eoconfuciusornis zhengi, IVPP V 11977; B. BMNHC-PH870, referred to E. zhengi in Navalón et al. (2018); C. C. sanctus. The arrowheads in B and C indicate that the deltopectoral crest is perforated by a suboval fenestra in BMNHC-PH870 and C. sanctus, but not in E. zhengi. Line drawings are not to scale

Fig. 12 Skeleton and anatomy of Confuciusornis sanctus [= 'Confuciusornis feducciai'], DNHM D2454 A. overview; B. skull in ventral view, showing that the mandibular symphysis is notched rostrally (arrow); C. furcula in ventral view; D. sternum in ventral view, showing the incomplete caudal end (dashed line) and a cleft at the midpoint of the rostral margin (arrow); E. pelvis in ventral viewScale bars equal 20 mm in A, 10 mm in B-E
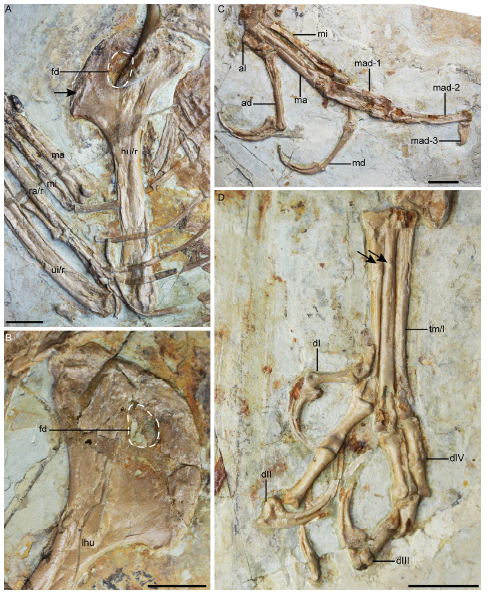
Fig. 13 Detail anatomy of Confuciusornis sanctus [= 'Confuciusornis feducciai'], DNHM D2454, showing that it is morphologically identical to C. sanctus A. right humerus in cranial view showing the ridge on the proximal part of the cranial surface of the deltopectoral crest (arrow); B. proximal left humerus in cranial view showing the perforated deltopectoral crest; C. dorsal view of metacarpals; D. dorsal view of metatarsals The arrows in D denote the dorsal tubercles on metatarsals II and III. Scale bars = 10 mm

Fig. 14 Skeleton and anatomy of Confuciusornis sanctus [='Confuciusornis jianchangensis'], PMOL-AB00114 A. overview; B. skull; C. thoracic vertebrae showing that the centra are excavated by lateral suboval fossae (arrow), as in C. sanctus; D. hindlimb. The arrows in D denote the dorsal tubercles on metatarsals II and III Scale bars = 20 mm
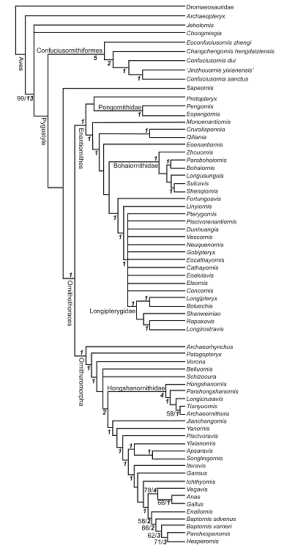
Fig. 15 Cladogram of Mesozoic birds showing interrelationships within the Confuciusornithiformes The strict consensus tree is built from 144 most parsimonious trees (Length = 1200; Consistency index = 0.324; Retention index = 0.673) that were found in the present phylogenetic analysis Bootstrap and Bremer values are indicated in bold and italic fonts near the corresponding nodes, respectively
| 1 |
Agnolín F L, Egli F B, Chatterjee S et al., 2017. Vegaviidae, a new clade of southern diving birds that survived the K/T boundary. Sci Nat, 104:87
DOI URL |
| 2 | Chiappe L M , 2002. Basal birds phylogeny: problems and solution. In: Chiappe L M, Witmer L M eds. Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. 448-472 |
| 3 | Chiappe L M, Meng Q J , 2016. Birds of Stone: Chinese Avian Fossils from the Age of Dinosaurs. Baltimore: Johns Hopkins University Press. 1-294 |
| 4 | Chiappe L M, Walker C A , 2002. Skeletal morphology and systematics of the Cretaceous Euenantiornithes (Ornithothoraces: Enantiornithes). In: Chiappe L M, Witmer L M eds. Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. 240-267 |
| 5 | Chiappe L M, Ji S A, Ji Q et al., 1999. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. Bull Am Mus Nat Hist, 242:1-89 |
| 6 |
Chiappe L M, Marugán-Lobón J, Ji S A et al., 2008. Life history of a basal bird: morphometrics of the Early Cretaceous Confuciusornis. Biol Lett, 4:719-723
DOI URL |
| 7 |
De Ricqlès A J, Padian K, Horner J R et al., 2003. Osteohistology of Confuciusornis sanctus (Theropoda: Aves). J Vert Paleont, 23:373-386
DOI URL |
| 8 |
Gao C L, Chiappe L M, Zhang F J et al., 2012. A subadult specimen of the Early Cretaceous bird Sapeornis chaoyangensis and a taxonomic reassessment of sapeornithids. J Vert Paleont, 32:1103-1112
DOI URL |
| 9 | Gao K Q, Li Q G, Wei M R et al., 2009. Early Cretaceous birds and pterosaurs from the Sinuiju Series, and geographic extension of the Jehol Biota into the Korean Peninsula. J Paleontol Soc Korea, 25:57-61 |
| 10 | Gill F B , 2007. Ornithology, 3 rd ed . New York: W. H. Freeman. 1-720 |
| 11 |
Goloboff P A, Catalano S A , 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics, 32:221-238
DOI URL |
| 12 | He H Y, Wang X L, Zhou Z H et al., 2004. Timing of the Jiufotang Formation (Jehol Group) in Liaoning, northeastern China, and its implications. Geophy Res Lett, 31:L12605 |
| 13 | He H Y, Wang X L, Jin F et al., 2006. The 40Ar/ 39Ar dating of the early Jehol Biota from Fengning, Hebei Province, northern China . Geochem Geophy Geosyst, 7:Q04001 |
| 14 | Hou L H , 1997. Mesozoic Birds of China. Nantou Country: Feng-Huang-Ku Bird Park of Taiwan. 1-228 |
| 15 | Hou L H, Zhou Z H, Gu Y C et al., 1995. Confuciusornis sanctus, a new Late Jurassic sauriurine bird from China. Chin Sci Bull, 40:1545-1551 |
| 16 |
Hou L H, Martin L D, Zhou Z H et al., 1999. A diapsid skull in a new species of the primitive bird Confuciusornis. Nature, 399:679-682
DOI URL |
| 17 | Hou L H, Zhou Z H, Zhang F C et al., 2002. Mesozoic Birds from Western Liaoning in China. Shenyang: Liaoning Science and Technology Publishing House. 1-120 |
| 18 |
Hu H, O’Connor J K , 2017. First species of Enantiornithes from Sihedang elucidates skeletal development in Early Cretaceous enantiornithines. J Syst Palaeontol, 15:909-926
DOI URL |
| 19 | Li D S , 2010. Osteology of Confuciusornis sanctus. MS thesis. Beijing: Graduate School of Chinese Academy of Sciences. 1-77 |
| 20 | Li L, Wang J Q, Hou S L , 2010. A new species of Confuciusornis from Lower Cretaceous of Jianchang, Liaoning, China. Glob Geol, 29:183-187 |
| 21 | Linnaeus C , 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Vol. 1: Regnum Animale. Editio decima, reformata. Stockholm: Laurentii Salvii. 1-824 |
| 22 |
Marugán-Lobón J, Chiappe L M, Ji S A et al., 2011. Quantitative patterns of morphological variation in the appendicular skeleton of the Early Cretaceous bird Confuciusornis. J Syst Palaeontol, 9:91-101
DOI URL |
| 23 |
Maxwell E E , 2008. Comparative embryonic development of the skeleton of the domestic turkey (Meleagris gallopavo) and other galliform birds. Zoology, 111:242-257
DOI PMID |
| 24 |
Navalón G, Meng Q J, Marugán-Lobón J et al., 2018. Diversity and evolution of the Confuciusornithidae: evidence from a new 131-million-year-old specimen from the Huajiying Formation in NE China. J Asian Earth Sci, 152:12-22
DOI URL |
| 25 |
O’Connor J K, Chiappe L M , 2011. A revision of enantiornithine (Aves: Ornithothoraces) skull morphology. J Syst Palaeontol, 9:135-157
DOI URL |
| 26 | O’Connor J K, Chiappe L M, Bell A , 2011. Pre-modern birds: avian divergences in the Mesozoic. In: Gareth J D, Gary K eds. Living Dinosaurs: the Evolutionary History of Birds. Oxford: Wiley. 39-114 |
| 27 | O’Connor J K, Wang M, Zheng X T et al., 2014. The histology of two female Early Cretaceous birds. Vert PalAsiat, 52:112-128 |
| 28 |
O’Connor J K, Zheng X T, Sullivan C et al., 2015. Evolution and functional significance of derived sternal ossification patterns in ornithothoracine birds. J Evol Biol, 28:1550-1567
DOI URL |
| 29 |
Pan Y H, Zheng W J, Moyer A E et al., 2016. Molecular evidence of keratin and melanosomes in feathers of the Early Cretaceous bird Eoconfuciusornis. Proc Natl Acad Sci USA, 113:E7900-E7907
DOI URL |
| 30 |
Sullivan C, Xu X , 2017. Morphological diversity and evolution of the jugal in dinosaurs. Anat Rec, 300:30-48
DOI URL |
| 31 |
Wang M, Hu H , 2017. A comparative morphological study of the jugal and quadratojugal in early birds and their dinosaurian relatives. Anat Rec, 300:62-75
DOI URL |
| 32 | Wang M, Zhou Z H , 2017a. The evolution of birds with implications from new fossil evidences. In: Maina J N ed. The Biology of the Avian Respiratory System: Evolution, Development, Structure and Function. Cham: Springer International Publishing. 1-26 |
| 33 |
Wang M, Zhou Z H , 2017b. A new adult specimen of the basalmost ornithuromorph bird Archaeorhynchus spathula (Aves: Ornithuromorpha) and its implications for early avian ontogeny. J Syst Palaeontol, 15:1-18
DOI URL |
| 34 | Wang M, Zhou Z H, O’Connor J K et al., 2014. A new diverse enantiornithine family (Bohaiornithidae fam. nov.) from the Lower Cretaceous of China with information from two new species. Vert PalAsiat, 52:31-76 |
| 35 |
Wang M, O’Connor J K, Pan Y H et al., 2017a. A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat Commun, 8:14141
DOI PMID |
| 36 |
Wang M, Li Z H, Zhou Z H , 2017b. Insight into the growth pattern and bone fusion of basal birds from an Early Cretaceous enantiornithine bird. Proc Natl Acad Sci USA, 114:11470-11475
DOI URL |
| 37 |
Wang Y, Hu H, O’Connor J K et al., 2017. A previously undescribed specimen reveals new information on the dentition of Sapeornis chaoyangensis. Cretaceous Res, 74:1-10
DOI URL |
| 38 | Zhang F C, Hou L H, Ou Y L , 1998. Osteological microstructure of Confuciusornis: preliminary report. Vert PalAsiat, 36:126-135 |
| 39 | Zhang F C, Zhou Z H, Benton M J , 2008. A primitive confuciusornithid bird from China and its implications for early avian flight. Sci China Ser A, 51:625-639 |
| 40 |
Zhang F C, Kearns S L, Orr P J et al., 2010. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature, 463:1075-1078
DOI URL |
| 41 |
Zhang Z H, Gao C L, Meng Q J et al., 2009. Diversification in an Early Cretaceous avian genus: evidence from a new species of Confuciusornis from China. J Ornithol, 150:783-790
DOI URL |
| 42 |
Zheng X T, Wang X L, O’Connor J K et al., 2012. Insight into the early evolution of the avian sternum from juvenile enantiornithines. Nat Commun, 3:1116
DOI URL |
| 43 |
Zheng X T, O’Connor J, Huchzermeyer F et al., 2013. Preservation of ovarian follicles reveals early evolution of avian reproductive behaviour. Nature, 495:507-511
DOI URL |
| 44 |
Zheng X T, O’Connor J K, Wang X L et al., 2017. Exceptional preservation of soft tissue in a new specimen of Eoconfuciusornis and its biological implications. Natl Sci Rev, 4:441-452
DOI URL |
| 45 |
Zhou S, Zhou Z H, O’Connor J K , 2013. Anatomy of the basal ornithuromorph bird Archaeorhynchus spathula from the Early Cretaceous of Liaoning, China. J Vert Paleont, 33:141-152
DOI URL |
| 46 |
Zhou Z H , 2014. The Jehol Biota, an Early Cretaceous terrestrial Lagerstätte: new discoveries and implications. Natl Sci Rev, 1:543-559
DOI URL |
| 47 |
Zhou Z H, Zhang F C , 2002. A long-tailed, seed-eating bird from the Early Cretaceous of China. Nature, 418:405-409
DOI URL |
| 48 |
Zhou Z H, Zhang F C , 2003. Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China. Can J Earth Sci, 40:731-747
DOI URL |
| 49 | Zhou Z H, Zhang F C , 2006a. Mesozoic birds of China: a synoptic review. Vert PalAsiat, 44:74-98 |
| 50 |
Zhou Z H, Zhang F C , 2006b. A beaked basal ornithurine bird (Aves, Ornithurae) from the Lower Cretaceous of China. Zool Scr, 35:363-373
DOI URL |
| 51 |
Zhou Z H, Zhang F C, Li Z H , 2010. A new Lower Cretaceous bird from China and tooth reduction in early avian evolution. Proc R Soc B, 277:219-227
DOI URL |
| [1] | 薛钦元, 余逸伦, 潘照晖, 朱幼安, 朱敏. 节甲鱼类(有颌脊椎动物干群)系统发育多样性的衰落与泥盆纪重大环境-生物事件耦合. 古脊椎动物学报, 2024, 62(1): 1-12. |
| [2] | 罗彦超, 朱敏, 卢立伍, 潘照晖. 湖南中泥盆统跳马涧组中华沟鳞鱼再研究. 古脊椎动物学报, 2023, 61(4): 261-276. |
| [3] | 冯东昊, 徐光辉, 马昕莹, 任艺. 云贵地区中三叠世近鲱形类贵州中华真颌鱼(Sinoeugnathus kueichowensis)的分类学修订. 古脊椎动物学报, 2023, 61(3): 161-181. |
| [4] | 王敏. 马氏副渤海鸟(鸟翼类:反鸟类)新标本对鸟类头骨早期演化的意义. 古脊椎动物学报, 2023, 61(2): 90-107. |
| [5] | 王世骐, 李春晓, 张晓晓. 乳齿象类的命名与中文译法及分类学问题讨论. 古脊椎动物学报, 2021, 59(4): 295-332. |
| [6] | 任艺, 徐光辉. 云南罗平中三叠世翼鳕属一新种及早期辐鳍鱼类系统发育关系. 古脊椎动物学报, 2021, 59(3): 169-199. |
| [7] | 李志恒, 王维, 胡晗, 王敏, 易鸿宇, 卢静. 眼齿鸟再分析显示其蜥蜴属性. 古脊椎动物学报, 2021, 59(2): 95-105. |
| [8] | 安晓青, 张兆群. 气候变化与早期兔形类的演化:基于内蒙古Ordolagus新材料的研究. 古脊椎动物学报, 2021, 59(2): 138-168. |
| [9] | 吴倩, 邹晶梅, 李志恒, 艾莉达. 现生鸟类和孔子鸟(Confuciusornis)叉骨软骨的研究:初步分析以及对中生代鸟类飞行方式的启示. 古脊椎动物学报, 2021, 59(2): 106-124. |
| [10] | 肖博, 盛桂莲, 袁俊霞, 王斯人, 胡家铭, 陈顺港, 姬海龙, 侯新东, 赖旭龙. 中国东北鹿亚科动物亚化石的古DNA分子鉴定及系统发育分析. 古脊椎动物学报, 2020, 58(4): 328-337. |
| [11] | 邹晶梅, 福雅曼, 王敏, 郑晓廷. 未成熟羽毛在早白垩世热河鸟类生物群幼年反鸟类中的首次报道. 古脊椎动物学报, 2020, 58(1): 24-44. |
| [12] | 刘玉海, 朱敏, 林翔鸿, 卢立伍, 盖志琨. 新疆塔里木盆地志留纪盔甲鱼类新知. 古脊椎动物学报, 2019, 57(4): 253-273. |
| [13] | 张驰. MrBayes分子钟定年之程序. 古脊椎动物学报, 2019, 57(3): 241-252. |
| [14] | 徐光辉, 马昕莹, 吴飞翔, 任艺. 云南中三叠世拱鱼目一新属种及早期铰齿鱼类系统发育关系再评估. 古脊椎动物学报, 2019, 57(3): 181-204. |
| [15] | 周亚纯, 舒柯文, 张福成. 中生代鸟类牙齿的退化及其可忽略的体重效应. 古脊椎动物学报, 2019, 57(1): 38-50. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||