
古脊椎动物学报 ›› 2020, Vol. 58 ›› Issue (2): 134-158.DOI: 10.19615/j.cnki.1000-3118.200310
王世骐1,2( ), 张晓晓1,2,3,4, 李春晓1,2,3
), 张晓晓1,2,3,4, 李春晓1,2,3
收稿日期:2019-09-26
出版日期:2020-04-20
发布日期:2020-04-20
作者简介:wangshiqi@ivpp.ac.cn
基金资助:
WANG Shi-Qi1,2(), ZHANG Xiao-Xiao1,2,3,4, LI Chun-Xiao1,2,3
Received:2019-09-26
Published:2020-04-20
Online:2020-04-20
摘要:
中国北方中中新世的乳齿象类戈壁锯齿象(Serridentinus gobiensis
中图分类号:
王世骐, 张晓晓, 李春晓. 戈壁锯齿象(Serridentinus gobiensis Osborn & Granger, 1932)和同心中新乳齿象(Miomastodon tongxinensis Chen, 1978)再研究:关于粗壮型轭齿象(Zygolophodon)的讨论. 古脊椎动物学报, 2020, 58(2): 134-158.
WANG Shi-Qi, ZHANG Xiao-Xiao, LI Chun-Xiao. Reappraisal of Serridentinus gobiensis Osborn & Granger and Miomastodon tongxinensis Chen: the validity of Miomastodon. Vertebrata Palasiatica, 2020, 58(2): 134-158.
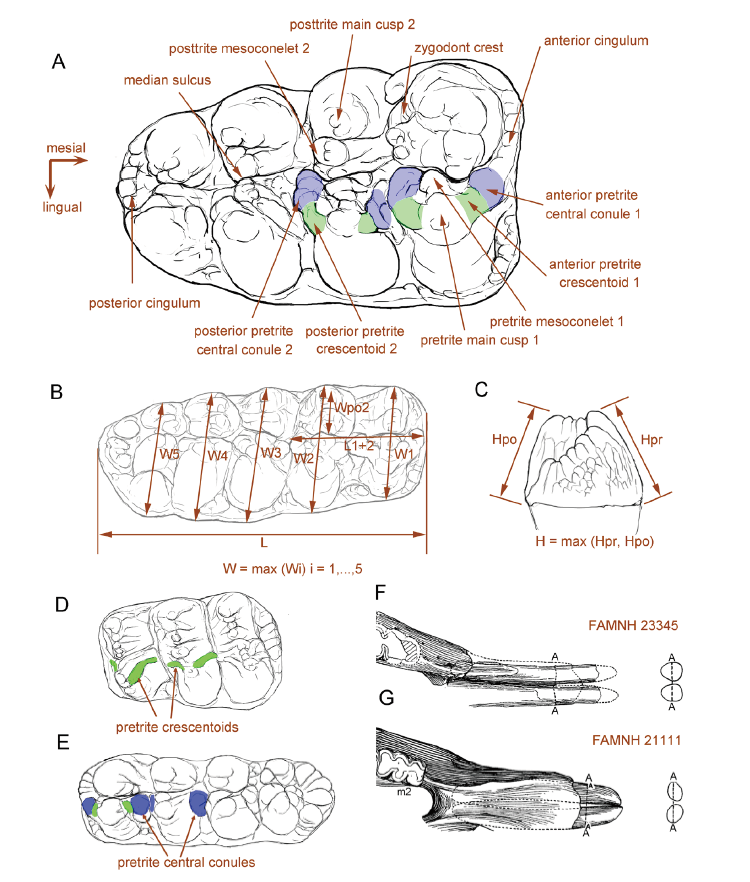
Fig. 1 Terminology and measurements of mastodont molars A. right M3 of Sinomastodon praeintermedius, denoting the terminology of tooth crown; B. molar crown measurements; C. molar height measurements; D, E. the development of pretrite accessory elements in a typical zygodont molar, Zygolophodon turicensis, only pretrite crescentoids present (D), and in a typical bunodont molar, Gomphotherium connexum, only pretrite central conule present in the lophid 2 (E);F, G. the cross-sections of lower tusks, showing the “erected oval cross-sectioned mandibular tusk” in Miomastodon merriami, in which Ddv > Dml (F), and the “laid oval cross-sectioned” in Gomphotherium productum, in which Ddv < Dml (G). Note that the discrimination of the pretrite accessory elements in the panels A, D, E: green color, pretrite crescentoids; blue color, pretrite central conules. Panels F and G were modified from Frick (1933). Abbreviations: L. length; L1+2. length of the first two loph(id)s; H. height;Hpo. height of the posttrite side; Hpr. height of the pretrite side; W. width;W1, 2, …, 5. width of the 1st, 2nd, …, 5th loph(id); Wpo2. width of the second posttrite half loph(id)
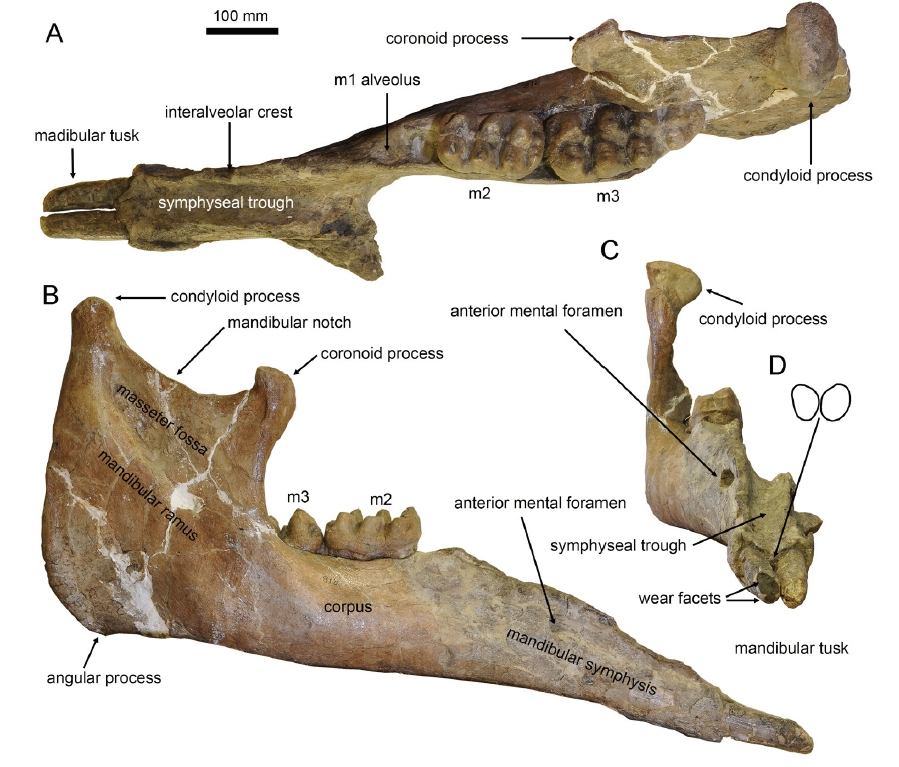
Fig. 2 Type mandible of Miomastodon gobiensis, AMNH 26461, from the Zhunwuguer locality, Tunggur region, Nei Mongol, in dorsal (A), lateral (B), and rostral (C) views, and the contour of the cross-section of mandibular tusks, in rostral view (D)
| maximum length | 1190 | |
|---|---|---|
| symphyseal length | 338.41 | |
| alveolar distance (from the most salient point of the retromolar trigonum to the symphyseal border of the corpus) | 424.1 | |
| ventral length measured from the gonion (angulus mandibular) to the tip of the symphysis | 965 | |
| width of corpus measured at the root of the ramus | 148.52 | |
| width of corpus measured at the anterioralveolus (or the grinding tooth if the alveolus is entirely resorbed) | 62.01 | |
| anterior symphyseal width | 113.75 | |
| maximum symphyseal width in the anterior part of the symphysis | 118.09 | |
| minimum symphyseal width in the posterior part of the symphysis | 69.88×2 | |
| maximum width of rostral trough in the anterior part of the symphysis | 80.17 | |
| minimum width of rostral trough in the posterior part of the symphysis | 82.76 | |
| rostral height measured at the symphyseal border (measurement taken perpendicular to the ventral border of the symphyseal rostrum) | 146.84 | |
| rostral height measured at the tip of rostrum (measurement as above) | 70.13 | |
| maximum mandibular height measured at the condyle perpendicular to the ventral border of the corpus | 467.97 | |
| maximum depth of ramus | 317.14 | |
| depth between gonion and coronoid processes | 364.98 | |
| height between gonion and condyle | 329.08 | |
| mid-alveolar length measured on the buccal side between the anterior alveolus (or grinding tooth if the alveolus is resorbed) and the root of the ramus | 321.73 | |
| exposed length of mandibular tusk | left | 134.25 |
| right | 131.5 | |
| width of mandibular tusk at the alveolu | left | 38.26 |
| right | 39.74 | |
| height of mandibular tusk at the alveolus | left | 45.60 |
| right | 47.32 | |
| length of wear facet of mandibular tusk | left | 35.33 |
| right | 48.31 |
Table 1 Mandibular and tusk measurements of Miomastodon gobiensis* (mm)
| maximum length | 1190 | |
|---|---|---|
| symphyseal length | 338.41 | |
| alveolar distance (from the most salient point of the retromolar trigonum to the symphyseal border of the corpus) | 424.1 | |
| ventral length measured from the gonion (angulus mandibular) to the tip of the symphysis | 965 | |
| width of corpus measured at the root of the ramus | 148.52 | |
| width of corpus measured at the anterioralveolus (or the grinding tooth if the alveolus is entirely resorbed) | 62.01 | |
| anterior symphyseal width | 113.75 | |
| maximum symphyseal width in the anterior part of the symphysis | 118.09 | |
| minimum symphyseal width in the posterior part of the symphysis | 69.88×2 | |
| maximum width of rostral trough in the anterior part of the symphysis | 80.17 | |
| minimum width of rostral trough in the posterior part of the symphysis | 82.76 | |
| rostral height measured at the symphyseal border (measurement taken perpendicular to the ventral border of the symphyseal rostrum) | 146.84 | |
| rostral height measured at the tip of rostrum (measurement as above) | 70.13 | |
| maximum mandibular height measured at the condyle perpendicular to the ventral border of the corpus | 467.97 | |
| maximum depth of ramus | 317.14 | |
| depth between gonion and coronoid processes | 364.98 | |
| height between gonion and condyle | 329.08 | |
| mid-alveolar length measured on the buccal side between the anterior alveolus (or grinding tooth if the alveolus is resorbed) and the root of the ramus | 321.73 | |
| exposed length of mandibular tusk | left | 134.25 |
| right | 131.5 | |
| width of mandibular tusk at the alveolu | left | 38.26 |
| right | 39.74 | |
| height of mandibular tusk at the alveolus | left | 45.60 |
| right | 47.32 | |
| length of wear facet of mandibular tusk | left | 35.33 |
| right | 48.31 |
| Miomastodon gobiensis | Miomastodon tongxinensis | |||||
|---|---|---|---|---|---|---|
| specimen | AMNH 26461 | AMNH 24676 | IVPP V 5584 | IVPP V 5585 | ||
| locus | right m2 | right m3 | right m3 | right m3 | right M3 | |
| L | 136.8 | 203.31 | 158.3 | 152.0 | 137.5 | |
| W1 | 75.19 | 91.29 | 74.65 | 69.0 | 81 | |
| W2 | 78.2 | ca.98 | 77.61 | 73.4 | 82 | |
| W3 | 87.52 | 75.51 | 67.6 | 66.5 | ||
| W4 | 63.34 | 51.5 | 44 | |||
| Hpo | 51.55+ | 68 | 42.89+ | 48.5+ | 48.5+ | |
Table 2 Cheek tooth measurements of Miomastodon* (mm)
| Miomastodon gobiensis | Miomastodon tongxinensis | |||||
|---|---|---|---|---|---|---|
| specimen | AMNH 26461 | AMNH 24676 | IVPP V 5584 | IVPP V 5585 | ||
| locus | right m2 | right m3 | right m3 | right m3 | right M3 | |
| L | 136.8 | 203.31 | 158.3 | 152.0 | 137.5 | |
| W1 | 75.19 | 91.29 | 74.65 | 69.0 | 81 | |
| W2 | 78.2 | ca.98 | 77.61 | 73.4 | 82 | |
| W3 | 87.52 | 75.51 | 67.6 | 66.5 | ||
| W4 | 63.34 | 51.5 | 44 | |||
| Hpo | 51.55+ | 68 | 42.89+ | 48.5+ | 48.5+ | |
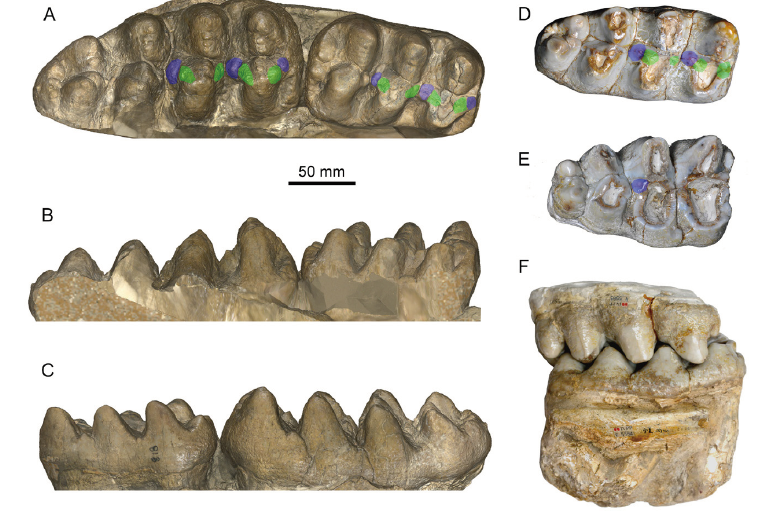
Fig. 3 Cheek teeth of Miomastodon gobiensis and Mio. tongxinensis A-C. Mio. gobiensis, AMNH 26461, right m2 and m3 of the type specimen, from the Zhunwuguer locality, Tunggur region, Nei Mongol, China, the ramus has been removed in the 3D digital model, in occlusal (A), buccal (B), and lingual (C) views; D-F. Mio. tongxinensis, from Tongxin region, Ningxia, China: IVPP V 5584, the type right m3, in occlusal view (D), V 5585, the referred right M3, in occlusal view (E), and the occlusion of V 5584 and V 5585, in buccal view (F), indicating that they belong to the same individual+Note: green color, pretrite crescentoids; blue color, pretrite central conules
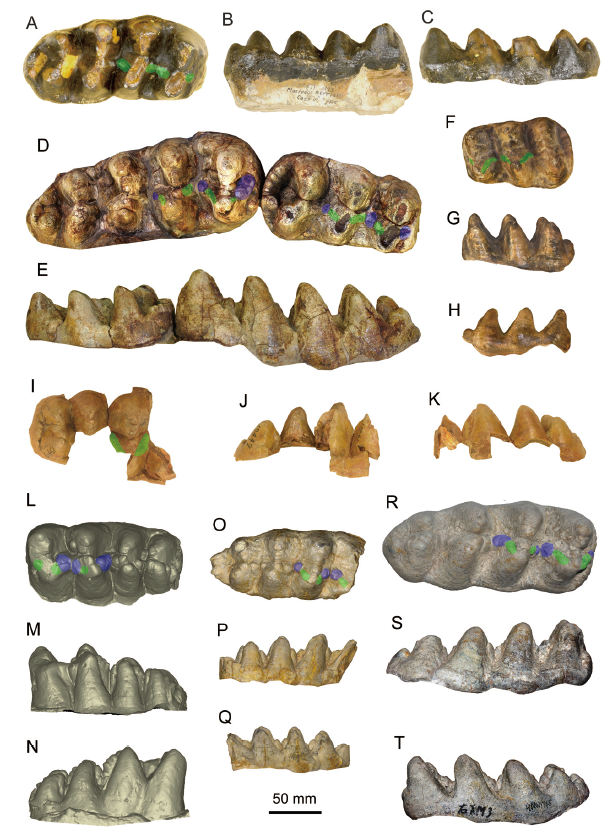
Fig. 4 Molars of the comparative species of the relevant taxa A-C. Miomastodon merriami, AMNH 14471 (type cast), the right m3, from Thousand Creek Formation, Nevada, USA, in occlusal (A), buccal (B), and lingual (C) views; D, E. Mio. metachinjiensis, AMNH 19414 (type specimen), the right m2 and m3, from Chinji Bungalow, Pakistan, in occlusal (D) and lingual (E) views; F-H. Zygolophodon turicensis, NMB OSM1288 (type cast), the left m2, from Elgg, Kanton Zürich, Switzerland, in occlusal (F), buccal (G), and lingual (H) views; I-K. Z. chinjiensis, AMNH 19447 (type specimen), the right M3, from Chinji Bungalow, Pakistan, in occlusal (I), lingual (J), and buccal (K) views; L-N. Gomphotherium productum, AMNH 14383 (type cast), the left m3, from Santa Fé, New Mexico, USA, in occlusal (L), buccal (M), and lingual (N) views; O-Q. G. subtapiroideum, SNSB-BSPG 1959 II 11381, the right m3, from Sandelzhausen, Germany, in occlusal (O), buccal (P), and lingual (Q) views; R-T. G. tassyi, HMV 1945, the right m3, from Linxia Basin, China, in occlusal (R), buccal (S), and lingual (T) views+Note: green color, pretrite crescentoids; blue color, pretrite central conules
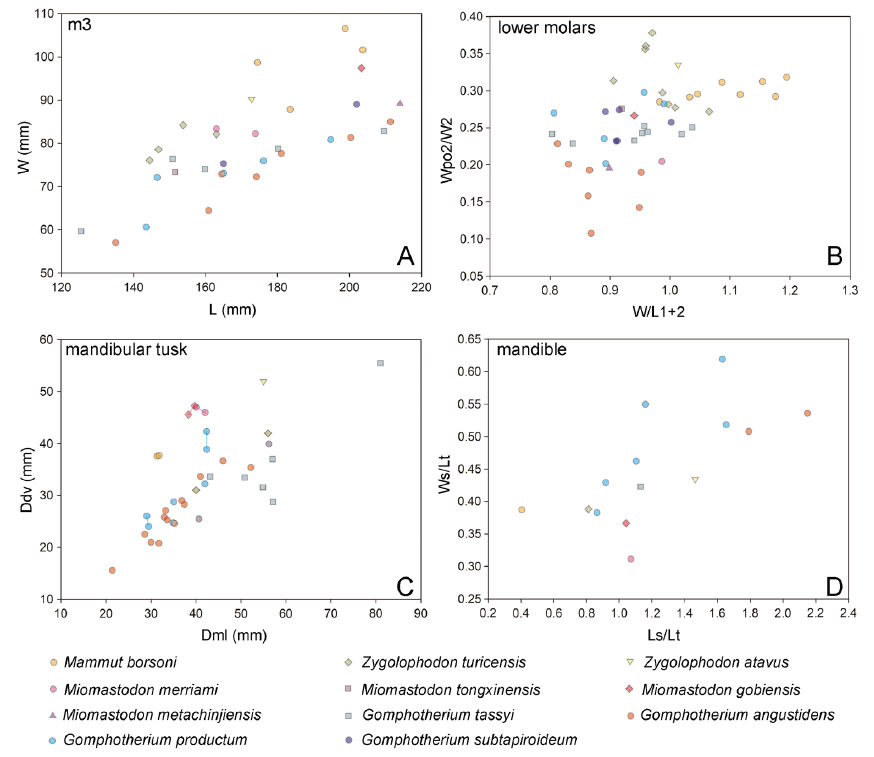
Fig. 5 Scatter plots of bunodont and zygodont cheek teeth and mandibular tusks of the relevant taxa A. m3 length vs. width (W/L); B. normalized maximal width (W/L1+2) vs. normalized posttrite width of lophid 2 (Wpo2/W2) for lower molars; C. cross-sectional mediolateral diameter (Dml) vs. dorsoventral (Ddv) diameter of mandibular tusk; D. mandibular measurements, the relative mandibular length (Ls/Lt) vs. the relative mandibular width (Ws/Lt). The raw data see Tables S1-S3
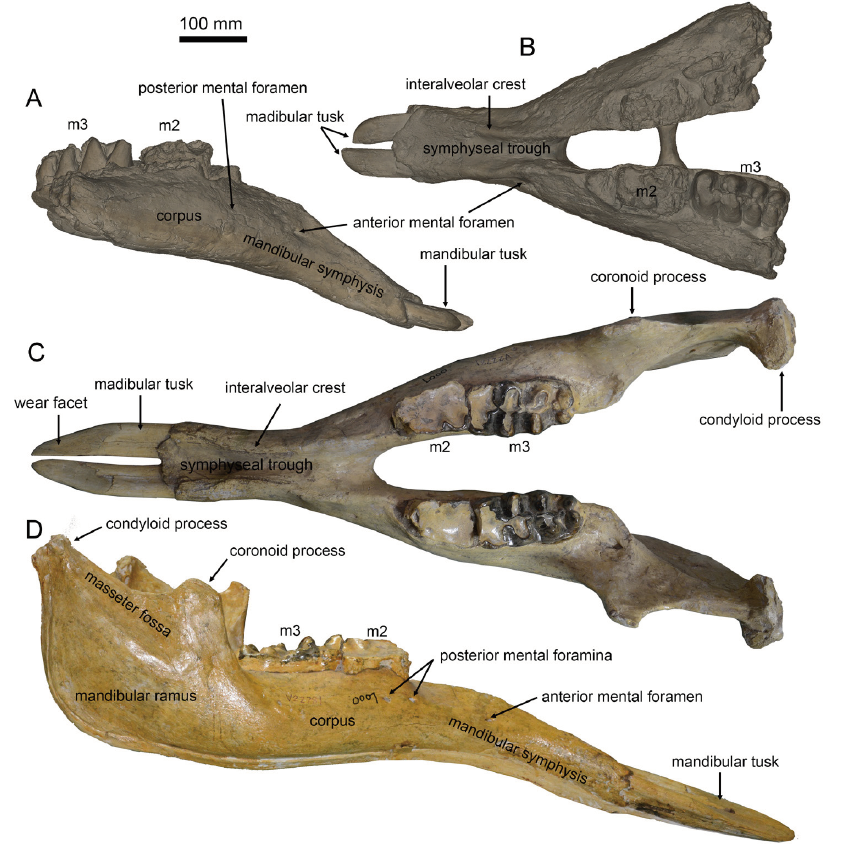
Fig. 6 Mandibles of Gomphotherium productum and G. tassyi, in comparison with Miomastodon gobiensis A, B. AMNH 14383, the type mandible cast of G. productum, from Santa Fé, New Mexico, USA, in lateral (A) and dorsal (B) views; C, D. IVPP V 22781, unpublished mandible of G. tassyi, from the Heijiagou locality, Zhongning region, in dorsal (C) and lateral (D) views
| Taxa | Zygolophodon turicensis | Miomastodon merriami | Miomastodon gobiensis | Gomphotherium productum complex | Gomphotherium subtapiroideum/tassyi |
|---|---|---|---|---|---|
| elongation of mandibular symphysis | moderately elongated | weakly elongated | moderately elongated | moderately elongated | moderately elongated |
| inclination of mandibular symphysis | slightly ventrally inclined | slightly ventrally inclined | slightly ventrally inclined | strongly ventrally inclined | strongly ventrally inclined |
| posterior mental foramina | ? | present | absent | present | present |
| cross-section of mandibular tusk | laid oval | erected oval | erected oval | laid oval to circular | pyriform |
| protrusion of mandibular tusk | short | short | short | short | long |
| enamel thickness | thin | thick | thick | thick | thick |
| mesiodistal distance of interlophids | wide | moderate | moderate | narrow | moderate |
| pretrite central conules | absent | absent or small | small and singular | large and serrate | small, singular or subdivided |
| posttrite half lophids | strongly subdivided | moderately subdivided | moderately subdivided | weakly subdivided or undivided | weakly subdivided or undivided |
| posttrite main cuspid and mesoconelet | separation clear or unclear | separation unclear | separation unclear | separation clear | separation clear |
| alternative position and chevron | absent | absent | present | absent | absent |
Table 3 Comparison of mandible and dentition among selected species of Zygolophodon, Miomastodon, and Gomphotherium
| Taxa | Zygolophodon turicensis | Miomastodon merriami | Miomastodon gobiensis | Gomphotherium productum complex | Gomphotherium subtapiroideum/tassyi |
|---|---|---|---|---|---|
| elongation of mandibular symphysis | moderately elongated | weakly elongated | moderately elongated | moderately elongated | moderately elongated |
| inclination of mandibular symphysis | slightly ventrally inclined | slightly ventrally inclined | slightly ventrally inclined | strongly ventrally inclined | strongly ventrally inclined |
| posterior mental foramina | ? | present | absent | present | present |
| cross-section of mandibular tusk | laid oval | erected oval | erected oval | laid oval to circular | pyriform |
| protrusion of mandibular tusk | short | short | short | short | long |
| enamel thickness | thin | thick | thick | thick | thick |
| mesiodistal distance of interlophids | wide | moderate | moderate | narrow | moderate |
| pretrite central conules | absent | absent or small | small and singular | large and serrate | small, singular or subdivided |
| posttrite half lophids | strongly subdivided | moderately subdivided | moderately subdivided | weakly subdivided or undivided | weakly subdivided or undivided |
| posttrite main cuspid and mesoconelet | separation clear or unclear | separation unclear | separation unclear | separation clear | separation clear |
| alternative position and chevron | absent | absent | present | absent | absent |
| [1] | Borissiak A A, 1936. Mastodon atavus n. sp., der primitivste vertreter der gruppe M. angustidens. Trav Inst Paleozool, Acad Sci URSS, 5:171-234 |
| [2] | Buckley M, Recabarren O P, Lawless C et al., 2019. A molecular phylogeny of the extinct South American gomphothere through collagen sequence analysis. Quat Sci Rev, 224:1-8 |
| [3] | Chen G F, 1978. Mastodont remains form the Miocene of Zhongning-Tongxin region in Ningxia. Vert PalAsiat, 16(2):103-110 |
| [4] | Chen G F, 1988. Mastodont remains from the Miocene of Junggar Basin in Xinjiang. Vert PalAsiat, 26(4):265-277 |
| [5] | Chow M C, Chang Y P, 1961. New mastodonts from North China. Vert PalAsiat, (3):245-255 |
| [6] | Chow M C, Chang Y P, 1974. Chinese Fossil Elephantoids. Beijing: Science Press. 1-74 |
| [7] | Chow M C, Chang Y P, You Y Z, 1978. Notes on some mastodons from Yunnan. Prof Pap Stratigr Palaeontol, 7:68-74 |
| [8] | Cope E D, 1873. Synopsis of New Vertebrata from the Tertiary of Colorado, Obtained During the Summer of 1873. Washington: Government Printing Office. 1-19 |
| [9] | Cope E D, 1875. On a new Mastodon and rodent. Proc Acad Nat Sci Philadelphia, 26:221-223 |
| [10] | Frick C, 1933. New remains of trilophodont-tetrabelodont mastodons. Bull Am Mus Nat Hist, 56:505-652 |
| [11] | Göhlich U B, 1998. Elephantoidea (Proboscidea, Mammalia) aus dem Mittel- und Obermiozän der oberen Süßwassermolasse Süddeutschlands: Odontologie und Osteologie. Münchner Geowiss Abh, 36:1-245 |
| [12] | Göhlich U B, 2010. The Proboscidea (Mammalia) from the Miocene of sandelzhausen (southern Germany). Paläontol Z, 84(1):163-204 |
| [13] | Hay O P, 1922. Further observations on some extinct elephants. Proc Biol Soc Wash, 35:97-101 |
| [14] | Illiger C D, 1811. Prodromus Systematis Mammalium et Avium Additis Terminis Zoographicis Uttriusque Classis. Berlin:Salfeld. 1-301 |
| [15] | Königswald W von, 2016. The diversity of mastication patterns in Neogene and Quaternary proboscideans. Palaeontogr Abt A: Paläozool, Stratigr, 307:1-41 |
| [16] | Lehmann U, 1950. Über Mastodontenreste in der Bayerischen Staatssammlung in München. Palaeontographica, 99(A):121-228 |
| [17] | Lofgren D L, Anand R S, 2011. Partial skull of Zygolophodon (Mammalia, Proboscidea) from the Barstow Formation of California. J Vert Paleont, 31(6):1392-1396 |
| [18] | Lortet L, Chantre E, 1878. Études Paléontologiques dans le Bassin du Rhone. Période Tertiaire. Recherches sur les Mastodontes et les Faunes mammalogiques qui les accompagnent. Arch Mus Hist Nat Lyon, 2:285-311 |
| [19] | Madden C T, Storer J E, 1985. The Proboscidea from the Middle Miocene Wood Mountain Formation, Saskatchewan. Can J Earth Sci, 22:1345-1350 |
| [20] | Osborn H F, 1921. First appearance of the true Mastodon in America. Am Mus Novit, 10:1-6 |
| [21] | Osborn H F, 1922. Dibelodon edensis (Frick) of southern California, Miomastodon of the Middle Miocene, new genus. Am Mus Novit, 49:1-4 |
| [22] | Osborn H F, 1923. New subfamily, generic, and specific stage in the evolution of the Proboscidea. Am Mus Novit, 99:1-4 |
| [23] | Osborn H F, 1929. New Eurasiatic and American proboscideans. Am Mus Novit, 393:1-28 |
| [24] | Osborn H F, 1936. Proboscidea: a Monograph of the Discovery, Evolution, Migration and Extinction of the Mastodonts and Elephants of the World. New York: The American Museum Press. 1-802 |
| [25] | Osborn H F, Granger W, 1932. Platybelodon grangeri, three growth stages, and a new Serridentine from Mongolia. Am Mus Novit, 537:1-13 |
| [26] | Qiu Z D, Wang X M, Li Q, et al., 2013. Neogene faunal succession and biochronology of central Nei Mongol (Inner Mongolia). In: Wang X M, Flynn L J, Fortelius M eds. Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology of Asia. New York: Columbia University Press. 155-186 |
| [27] | Schlesinger G, 1917. Die Mastodonten des K. K. Naturhistorischen Hofmuseums. Denkschr K K Naturhist Hofm, Geol-pal äont, 1:1-231 |
| [28] | Schneider C A, Rasband W S, Eliceiri K W, 2012. NIH Image to ImageJ: 25 years of image analysis. Nat Meth, 9:671-675 |
| [29] | Spock L E, 1929. Pliocene beds of the Iren Gobi. Am Mus Novit, 394:1-8 |
| [30] | Tassy P, 1977. Découverte de Zygolophodon turicensis (Schinz) (Proboscidea, Mammalia) au lieu-dit Malartic a Simorre, Gers (Vindobonien moyen): implications paléoécologiques et biostratigraphiques. Géobios, 10(5):655-669 |
| [31] | Tassy P, 1983. Les Elephantoidea Miocènes du Plateau du Potwar, Groups de Siwalik, Pakistan. Ire Partie: Cadre chronologique et géographique, Mammutidés, Amébélodontidés. Ann Paléontol, 69(2):99-136 |
| [32] | Tassy P, 1985. La place des mastodontes Miocènes de l’ancien monde dans la phylogénie des Proboscidea (Mammalia): hypothèses et conjectures. Paris: Thèse Doctorat ès Sciences, UPMC(Université Pierre et Marie CURIE). 1-861 |
| [33] | Tassy P, 2013. L'anatomie cranio-mandibulaire de Gomphotherium angustidens (Cuvier, 1817) (Proboscidea, Mammalia): données issues du gisement d'En Péjouan (Miocène moyen du Gers, France). Geodiversitas, 35(2):377-445 |
| [34] | Tassy P, 2014. L'odontologie de Gomphotherium angustidens (Cuvier, 1817) (Proboscidea, Mammalia): données issues du gisement d'En Péjouan (Miocène moyen du Gers, France). Geodiversitas, 36(1):35-115 |
| [35] | Tobien H, 1972. Status of the genus Serridentinus Osborn 1923 (Proboscidea, Mammalia) and related forms. Mainzer Geowiss Mitt, 1:143-191 |
| [36] | Tobien H, 1975. The structure of the mastodont molar (Proboscidea, Mammalia). Part 2: the zygodont and the zygobunodont patterns. Mainzer Geowiss Mitt, 4:195-233 |
| [37] | Tobien H, 1996. Evolution of zygodons with emphasis on dentition. In: Shoshani J, Tassy P eds. The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives. Oxford: Oxford University Press. 76-88 |
| [38] | Tobien H, Chen G F, Li Y Q, 1988. Mastodonts (Proboscidea, Mammalia) from the late Neogene and Early Pleistocene of the People’s Republic of China, part II: the genera Tetralophodon, Anancus, Stegotetrabelodon, Zygolophodon, Mammut, Stegolophodon. Mainzer Geowiss Mitt, 17:95-220 |
| [39] | Wang S Q, Li Y, Duangkrayom J et al., 2017. A new species of Gomphotherium (Proboscidea, Mammalia) from China and the evolution of Gomphotherium in Eurasia. J Vert Paleont, 37(3):1-15 |
| [40] | Wang X M, Qiu Z D, Opdyke N D, 2003. Litho-, bio-, and magnetostratigraphy and paleoenvironment of Tunggur Formation (Middle Miocene) in central Inner Mongolia, China. Am Mus Novit, 3411:1-31 |
| [1] | 李世杰, 邓涛. 河北磁县九龙口中中新世动物群中真犀的再研究. 古脊椎动物学报, 2023, 61(3): 198-211. |
| [2] | 杨湘雯,李 雨, 王世骐 . 临夏盆地中中新统维曼嵌齿象(Gomphotherium wimani) (长鼻目,嵌齿象科)的头骨及颊齿. 古脊椎动物学报, 2017, 55(4): 331-346. |
| [3] | 邓涛,卢小康,史勤勤,孙博阳,王世骐. 甘肃临夏盆地中中新世皇冠鹿一新种. 古脊椎动物学报, 2014, 52(2): 170-182. |
| [4] | 陈少坤,刘 艳 . 河北磁县九龙口中中新世“Macrotherium cf. M. brevirostris”的分类地位. 古脊椎动物学报, 2013, 51(3): 205-210. |
| [5] | 王世骐,刘善品,颉光普,刘 佳,彭廷江,侯素宽. 甘肃天水武山县南峪村的维曼嵌齿象及其生物地层学意义. 古脊椎动物学报, 2013, 51(1): 71-84. |
| [6] | 吴文裕,倪喜军,叶 捷,孟 津,毕顺东. 新疆准噶尔盆地北缘中中新世早期的原圆齿鼠(Promylagaulinae, Mylagaulidae). 古脊椎动物学报, 2013, 51(1): 55-70. |
| [7] | 张兆群,Anu Kaakinen, 王李花,刘丽萍,刘 艳,傅铭楷. 内蒙古大庙中中新世上猿化石地点的鼠兔科化石. 古脊椎动物学报, 2012, 50(3): 281-292. |
| [8] | Sevket SEN, Margarita A. ERBAJEVA. 贝加尔湖区中中新世Aya洞穴地点戈壁古仓鼠一新种. 古脊椎动物学报, 2011, 49(3): 257-274. |
| [9] | 魏涌澎. 新疆准噶尔盆地北缘中中新世阿特拉旱松鼠及其生态环境讨论. 古脊椎动物学报, 2010, 48(3): 220-234. |
| [10] | Benjamin H. PASSEY , Jussi T. ERONEN , Mikael FORTELIUS , 张兆群. 华北晚中新世羚羊的食性与古环境——稳定碳同位素的证据. 古脊椎动物学报, 2007, 45(2): 118-127. |
| [11] | 叶捷, 吴文裕, 孟津. 准噶尔盆地北缘哈拉玛盖组中的Anchitherium. 古脊椎动物学报, 2005, 43(02): 100-109. |
| [12] | 伍少远. Alloptox gobiensis (兔形目,鼠兔科) 头骨形态及系统位置. 古脊椎动物学报, 2003, 41(02): 115-130. |
| [13] | 吴文裕, 孟 津, 叶 捷. 新疆准噶尔盆地北缘Pliopithecus的发现. 古脊椎动物学报, 2003, 41(01): 76-86. |
| [14] | 魏明瑞, 郭建崴. 宁夏同心中中新世三种植食性哺乳动物牙齿碳同位素分析. 古脊椎动物学报, 2002, 40(04): 300-304. |
| [15] | 吴文裕, 叶,捷, 孟津 毕顺东, 刘丽萍 , 张翼. 新疆准噶尔盆地北缘中中新世兔形类新材料. 古脊椎动物学报, 1998, 36(04): 319-329. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||